
The 37 species and 8 non-nominotypical subspecies of Ips DeGeer, 1775 bark beetles of the World are included in a first comprehensive key to adults. Males and females of species with diagnostically important dimorphism are treated separately. Diagnostic morphological summaries are presented for all species and subspecies in fact sheets. Fact sheets include over 200 illustrations, with rotatable images of species that are little known or economically significant pests (I. amitinus (Eichhoff, 1872), I. calligraphus (Germar, 1824), I. cembrae (Heer, 1836), I. confusus (LeConte, 1876), I. duplicatus (Sahlberg, 1836), I. grandicollis (Eichhoff, 1868), I. hauseri Reitter, 1894, I. lecontei Swaine, 1924, I. perturbatus (Eichhoff, 1869), I. pini (Say, 1826), I. sexdentatus (Boerner, 1767), I. shangrila Cognato and Sun, 2007, I. stebbingi Strohmeyer, 1908, I. subelongatus (Motschulsky, 1860), I. typographus (Linnaeus, 1758), I. woodi Thatcher, 1965). Links to an online peer-reviewed LUCID key to world Ips species and downloadable LUCID files are provided.

Ips pini (Photo by K. Bolte)
Ips bark beetles (Coleoptera: Curculionidae: Scolytinae: Ipini), are sub-cortical phloem feeders in Pinaceae (conifer trees), especially Pinus (pine), Picea (spruce) and Larix (larch or tamarack) species (Cognato, 2015). In non-outbreak times, Ips beetles mainly inhabit weakened or dead trees (Cognato, 2015). Adults and larvae kill healthy trees during outbreaks (Cognato, 2015) by destroying the phloem and cambium in tree trunks and limbs when feeding and tunnelling (Furniss and Carolin, 1977). Outbreaks can destroy thousands of hectares of healthy trees (Cognato, 2015). Some or all Ips bark beetles also transmit pathogenic fungi (Krokene and Solheim, 1998; Meng et al., 2015), in particular blue stain fungi (genera Grosmannia and Ceratocystis, Ascomycota: Sordariomycetes). Ceratocystis fungifrom Ips beetlesalso interfere with biological control of the conifer pest Sirex noctilio Fabricius (Hymenoptera: Siricidae) (Yousuf et al., 2014). Certain climatic conditions (e.g. droughts) may promote Ips outbreaks (Wermelinger, 2004; Breshears et al., 2005; Marini et al., 2017). Trees injured in outbreaks are sometimes later killed by Dendroctonus Erichson, 1836 bark beetles (Furniss and Carolin, 1977).Native Ips species are present in all countries where Pinus and Picea occur naturally (Cognato, 2015). Five Ips species (I. apache (full names presented in Table 1), I. calligraphus, I. grandicollis, I. subelongatus, and I. typographus) also occur as introduced species, especially in non-native plantations of Picea and Pinus. Some Ips species use Larix as a primary host tree genus (Table 1). A few species use Abies (fir) and Cedrus (true cedar) as hosts during outbreaks (Wood and Bright, 1992). Pseudostuga may be attacked occasionally outside its natural range (e.g. by Ips acuminatus).
There are 37 valid Ips species worldwide (Table 1). Phylogenetic analyses of the Ipini prompted transfer of several species to the genera Pseudips (Cognato, 2000) and Orthotomicus (Cognato and Vogler, 2001). Cognato (2015) reviewed literature on phylogeny, taxonomy, diagnosis and biology of all Ips species.
The genus Ips can be recognizedand identified to species by adult external morphology. Descriptions and regional keys to the species of Ips based on morphology are available (Balachowsky, 1949; Kurentsov and Kononov, 1966; Bright 1976; Grüne, 1979; Schedl, 1981; Wood, 1982; Holzschuh, 1988; Lanier etal., 1991; Pfeffer, 1995; Cognato and Sun, 2007). Cognato (2015) provides diagnoses for all world species. A generic key to Scolytinae larvae of eastern Canada is available (Thomas, 1957) but juvenile stages cannot be used for reliable identification on a global scale. Although Ips species have been discovered and identified using DNA sequence data (Cognato and Sun, 2007), validated protocols for universal DNA identification of Ips species have not yet been developed (Chang et al., 2012). Additional work is needed to demonstrate that DNA sequence records provide accurate identifications.The following tribal-level diagnostic characters for Ipini are modified from Wood (1986): compound eye sinuate (narrowed at mid-height), ventral half narrower than dorsal part; antennal scape (basal segment) slender elongate, funicle 5-segmented, club either obliquely truncate or sutures on posterior face strongly displaced toward apex; pronotum strongly declivous on anterior half (posterior half approximately horizontal, anterior half descends abruptly), with large asperities (broad spines); procoxae contiguous, intercoxal piece deeply notched or absent; protibia with three or four socketed denticles; scutellar shield visible in dorsal view; elytral declivity moderately sulcate to strongly excavated, sides with tubercles or spines in most; vestiture hair-like (not scale-like or wider at midlength than at base), except for branched hairs at anterior opening of prothorax.Ips can be separated from other genera of Ipini by the combined features of the antennal club and elytral declivity. The following diagnostic characters are modified from Wood (1986), as done by Cognato (2000) and Cognato and Vogler (2001):
Ips is most similar in appearance to two other Ipini genera that also develop in Pinaceae: Orthotomicus Ferrari, 1867 and Pseudips Cognato, 2000. Ips can be distinguished from Orthotomicus by the acute spine 2 of its elytral declivity (right-angled in many Orthotomicus) and the broader explanate edge of its elytral declivity. Ips can be distinguished from Pseudips by its straight, bisinuate or acutely angulate antennal club sutures. These sutures are broadly procurved (curved away from the antennal base at the midline of the club) in Pseudips, and also in the tropical, angiosperm feeding Acanthotomicus Blandford, 1894 and the warm-climate, ambrosia-feeding Premnobius Eichhoff, 1878. Pityogenes Bedel, 1888 and Pityokteines Fuchs, 1911 are conifer-feeding Ipini, recognized by their small size (1.8–3.7 mm) and the rounded edges of their elytral declivity. The tropical, ambrosia fungus feeding Premnophilus Brown, 1962 lacks visible antennal sutures.Most Ips species are grouped into subgenera, based on phylogenetic results by Cognato and Vogler (2001) and Cognato and Sun (2007). Diagnostic characteristics (external morphology only) of subgenera are as follows: Cumatotomicus Ferrari, body length >5 mm, spines on first and second elytral interstriae on declivity; Bonips Cognato, elytral declivity with four spines per side, elytral disc without punctures on interstriae; Granips Cognato, elytral declivity with five to six spines per side; Ips DeGeer, elytral declivity with four spines per side, elytral disc with punctures on interstriae; Incertae sedis, several Ips species are outside any named subgenus.Diagnostic characters of Ips spp. adults are based on Cognato (2015) and characters developed here. The closely-related (Cognato and Sun, 2007) species: I. confusus and I. paraconfusus, and also I. cembrae and I. subelongatus, and some individuals or one sex of several other species are not fully distinguished from each other in the key to species. This missing information may be important as these species may differ in their biology and distribution and whether they are a regulated pest or not (Stauffer et al., 2001). Additional examination by Ips specialists with appropriate reference collections is required to identify these specimens to species level using morphology (Cognato, 2015). DNA-based studies have been published to support identification of I. confusus and I. paraconfusus (Cognato et al., 1995; Cognato and Sun, 2007) and I. cembrae and I. subelongatus (Stauffer et al., 2001; Cognato and Sun, 2007) but these results have not yet been developed into identification tools.Ips species are distinguished primarily by characters of the elytra and frons. Identifiers usually begin by counting spines of the elytral declivity (i.e., along one edge of the apical elytral concavity). The following elytral declivity characters are useful: the number of spines on the declivity (not including small denticles on the first elytral interstria); the distance from spine 1 to the elytral suture relative to its height or to its distance from spine 2; and the shininess of the declivity compared to the elytral dorsal surface. Several characters come from the third declivital spine: its shape (acute, right-angled, and obtuse or rounded) and its profile (simple triangular); straight sided with acute apex; petiolate (narrower near base than near apex); hooked (with second point on ventral side); double pointed (appearing like two basally fused spines)). On the elytral disc (the horizontal part of the elytra), the presence or absence of punctures on the interstriae (elevated smooth surfaces between striae) are important, especially on the second and third interstriae midway between the anterior edge of the elytra and the declivity.
On the frons the following presence or absence characters are used: of a median tubercle; of a median carina (between median tubercle and labrum if both present); of a median fossa or pit (above median tubercle if present); of scattered circular tubercles; of setae; of dense setal brushes obscuring integument; or of setal punctures. A few species pairs can only be distinguished by the number of ridges on the pars stridens (Lanier et al., 1991), a stridulatory organ at the vertex of the head capsule. However, this technique is not included here because it requires removal of the head.
Specimens were examined from the Canadian National Collection of Insects, Arachnids and Nematodes (CNC, Ottawa, Canada), Michigan State University: A.J. Cook Arthropod Research Collection (MSUC, E. Lansing, Michigan, USA), and the collection of Miloš Knížek, Prague, Czech Republic. Specimens used to generate the key were previously identified by A. Cognato, D. Bright, and M. Knížek. Twenty to 100 specimens per species were examined for all species except for the following, for which only 1 to 10 specimens were available: I. chinensis, I. hauseri, I. longifolia, I. pilifrons thatcheri, I. schmutzenhoferi, I. shangrila, I. stebbingi, I. subelongatus. Sarah Smith and the USA National Museum of Natural History, Smithsonian Institution, Washington, D.C. provided images of the holotype of I. pilifrons thatcheri. Specimens were photographed using a Leica M205c stereoscope and multifocus images assembled using Leica LAS 4.8 software. Pale specimens were favoured for photography to increase visual contrast of spines and punctures against background integument. The key and species descriptions were built using LUCID 3.6 software. Primary type specimens were not examined, so morphological summaries in associated fact sheets should be considered as diagnostic tools only.
Measurements: elytral disc punctures are measured across the steepest part of the puncture walls on the midlength of the flatter anterior half elytra on interstriae 2 and 3; interstriae (also on disc) are bounded by the steepest parts of adjacent strial punctures. Measurement values presented in parentheses are considered outliers, showing rare variations of size or proportion. Males and females were treated separately in keys and fact sheets where external morphology of sexes was found to differ.
Table 1. Worldwide list of Ips species and subspecies with distribution and principal host genera (from Cognato, 2015). PDF fact sheets for some species include rotatable multi-angle habitus images.
| Species | Authority | Native Distribution* | Principal host | Fact Sheet |
| Ips acuminatus | (Gyllenhal, 1827) | Eurasia | Pinus | |
| Ips amitinus | (Eichhoff, 1872) | Eurasia (west) | Picea, Pinus | |
| Ips apache | Lanier, 1991 | North America (south) | Pinus | |
| Ips avulsus | (Eichhoff, 1868) | North America (east) | Pinus | |
| Ips bonanseai | (Hopkins, 1905) | North America (south) | Pinus | |
| Ips borealis borealis | Swaine, 1911 | North America (north) | Picea | |
| Ips borealis lanieri | Wood, 1974 | North America (north) | Picea | |
| Ips borealis swainei | Hopping, 1939 | North America (north) | Picea | |
| Ips borealis thomasi | Hopping, 1965 | North America (north) | Picea | |
| Ips calligraphus | (Germar, 1824) | North America | Pinus | |
| Ips cembrae | (Heer, 1836) | Eurasia (widespread) | Larix | |
| Ips chinensis | Kurentsov and Kononov, 1966 | Eurasia (southeast) | Pinus | |
| Ips confusus | (LeConte, 1876) | North America (west) | Pinus | |
| Ips cribricollis | (Eichhoff, 1868) | North America (south) | Pinus | |
| Ips duplicatus | (Sahlberg, 1836) | Eurasia (widespread) | Picea | |
| Ips emarginatus | (LeConte, 1876) | North America (west) | Pinus | |
| Ips grandicollis | (Eichhoff, 1868) | North America (east, south) | Pinus | |
| Ips hauseri | Reitter, 1894 | Eurasia (central) | Picea | |
| Ips hoppingi | Lanier, 1970 | North America (southwest) | Pinus | |
| Ips hunteri | Swaine, 1917 | North America (west) | Picea | |
| Ips integer | (Eichhoff, 1869) | North America (west, south) | Pinus | |
| Ips knausi | Swaine, 1915 | North America (west) | Pinus | |
| Ips lecontei | Swaine, 1924 | North America (south) | Pinus | |
| Ips longifolia | (Stebbing, 1909) | Eurasia (central) | Pinus | |
| Ips montanus | (Eichhoff, 1881) | North America (west) | Pinus | |
| Ips nitidus | Eggers, 1933 | China | Picea | |
| Ips paraconfusus | Lanier, 1970 | North America (west) | Pinus | |
| Ips perroti | Swaine, 1915 | North America (north) | Pinus | |
| Ips perturbatus | (Eichhoff, 1869) | North America (north) | Picea | |
| Ips pilifrons pilifrons | Swaine, 1912 | North America (west) | Picea | |
| Ips pilifrons sulcifrons | Wood, 1960 | North America (west) | Picea | |
| Ips pilifrons thatcheri | Wood, 1975 | North America (west) | Picea | |
| Ips pilifrons utahensis | Wood, 1960 | North America (west) | Picea | |
| Ips pini | (Say, 1826) | North America (widespread) | Pinus | |
| Ips plastographus maritimus | Lanier, 1970 | North America (west) | Pinus | |
| Ips plastographus plastographus | (LeConte, 1868) | North America (west) | Pinus | |
| Ips schmutzenhoferi | Holzschuh, 1988 | Asia (Himalayas) | Larix, Picea, Pinus | |
| Ips sexdentatus | (Boerner, 1767) | Eurasia (widespread) | Pinus | |
| Ips shangrila | Cognato and Sun, 2007 | Asia (east) | Picea | |
| Ips stebbingi | Strohmeyer, 1908 | Eurasia (central) | Picea, Pinus | |
| Ips subelongatus | (Motschulsky, 1860) | Eurasia (east) | Larix | |
| Ips tridens engelmanni | Swaine, 1917 | North America (west) | Picea | |
| Ips tridens tridens | (Mannerheim, 1852) | North America (west) | Picea | |
| Ips typographus | (Linnaeus, 1758) | Eurasia (north and west) | Picea | |
| Ips woodi | Thatcher, 1965 | North America (west) | Pinus |
* South = tropical and subtropical parts of North America. North America refers to the North American continent including all countries north and west of Colombia. Widespread may not include all countries in the continent.
The illustrated dichotomous key presented here and interactive LUCID keys developed both include multiple routes to identification of one or both sexes of many species at the following internet URL: https://keys.lucidcentral.org/keys/v3/ips_de_geer/.
In some cases multiple identification paths were necessary because of observed intraspecific variation. In other cases species were scored as having additional character states because observed morphology was too close to the dividing line between character states to assign specimens reliably to one state or the other. Although these divergent interpretations of morphology sometimes contradict published descriptions they do not necessarily imply that those descriptions are incorrect. Identification of some Ips specimens remains difficult despite these steps. Users should compare identifications carefully to the fact sheet for that species or other taxonomic literature where indicated to maximize identification quality. Both keys are intended for use with single specimens (or partial specimens) of unknown origin for use with samples from traps or international plant quarantine interceptions. However, morphological information from both sexes, tree host species and geographic origin can all help improve accuracy of identifications.
Subspecies included in the key are defined partly based on morphology and partly on geography. Their taxonomic validity and limits remain under investigation. The morphological key to species may not produce accurate subspecies-level identifications when used without added locality-based inference. Entries were made for Orthotomicus and Pseudips using characters from multiple species to assist users who may have confused members of these genera for Ips species.
Head A B C D E F GHIJKLMNOPQR S T UVWXYZ
Elytra ABC D E FG H I JKLMNO P QR S T UVWXYZ
Other ABCDEF G HIJKLMNO P QR S TUVWXYZ
angulate: Antennal sutures with angulate peaks and valleys.

antennal club sutures:
Lines dividing antennal club into three antnnomeres.
Figure shows two angulate sutures near middle of oval club.

bisinuate:
Antennal sutures curved through multiple peaks and valleys.

central carina:
Ridge or elongate tubercle at midline of frons (circled). Not part of epistomal tubercles.

epistomal:
Epistoma. Area of head immediately above mouth opening. The epistomal row of tubercles (circled) is immediately above the epistoma. Epistomal setae project downward from the epistoma.

fovea:
A depression or shallow pit.
frons:
Portion of head between eyes. Extends vertically from epistoma to top of eyes. Ips pini, male frons
stridulatory apparatus:
Rows of parallel ridges at the posterior end of the female head (or pars stridens, arrow) and on the inner surface of the pronotum used for stridulatory sound production. Observing these structures requires removal of the head. See Lanier et al., 1991 for details about using this character system. Ips pini dorsal view of female head showing pars stridens (normally hidden under pronotum)
transverse carina:
Ridge or elongate tubercle across midline of frons. Impunctate in some species, weakly raised and impunctate in other species.
Ips sexdentatus female head showing transverse carina
declivital apex:
Posterior-most part of elytra when elytra are closed (bottom of figure). Also the apical-most part of the elytra.
declivity:
The apical (posterior) sloping part of the elytra. In Ips the declivity is concave with spines along the sides.
disc:
The basal (anterior) level part of elytra. Does not include laterally sloped interstriae 5 to 9, or declivity. Ips confusus, structures
elytron:
One of the hardened anterior pair of wings. These together form the elytra, the longitudinally divided dorsal shell covering the posterior half of the beetle.
emarginate:
Declivital spine with two apices (like two spines fused at base).
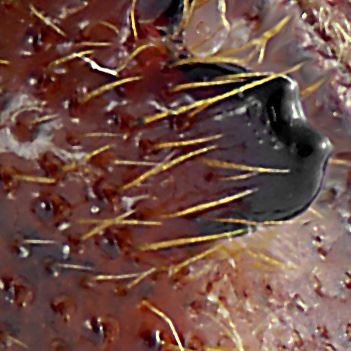
hooked:
Declivital spine with point at both apex and ventral side.

impunctate:
Without puncture-like depressions on the interstriae (here seen as broad pale transverse bands). Measured on interstriae 2 and 3 on middle third of elytra (box).

interstriae:
The area between major rows of punctures on elytra (arrow). In this image these appear as shining elevated horizontal stripes. One or more interstriae bear interstrial punctures in some species. Interstriae are counted sequentially from the midline of the beetle beginning between the suture and the first stria. White rectangle shows area of elytral disc used for comparison of species.

Ips montanus, with box showing area examined for interstrial punctures.
interstrial punctures:
Punctures on interstriae. Presence or absence of interstrial punctures is assessed on middle part of elytral disc. Interstrial punctures bear setae unless setae are broken off.
Ips montanus, with box showing area examined for interstrial punctures.
parallel sided:
Declivital spine nearly parallel-sided on basal half, and strongly tapered at apex.

petiolate:
Declivital spine narrowed near base.
Synonyms = pedunculate, capitate.

punctate:
With puncture-like depressions in integument. These bear setae (hairlike projections) unless the setae are broken off.

spine:
A horn-like outgrowth of the integument that is not separated by any groove or ridge. Spines are present on the elytral declivity of all Ips species.
striae:
The largest rows of punctures on the elytra. In Ips these are interrupted or distorted at the declivity. Striae are counted sequentially from the midline of the beetle.

strial punctures:
Punctures along the striae. Larger than interstrial punctures and positioned on strial groove.

suture:
The elytral suture is the line (or gap) at the midline of the beetle where the two elytra meet.
tapered:
Declivital spine narrowed evenly to apex.
Synonym = acuminate.

gallery: A connected system of subcortical tunnels and chambers in which a group of Ips beetles reproduce and develop. Galleries are usually initiated by a male and one or more females.
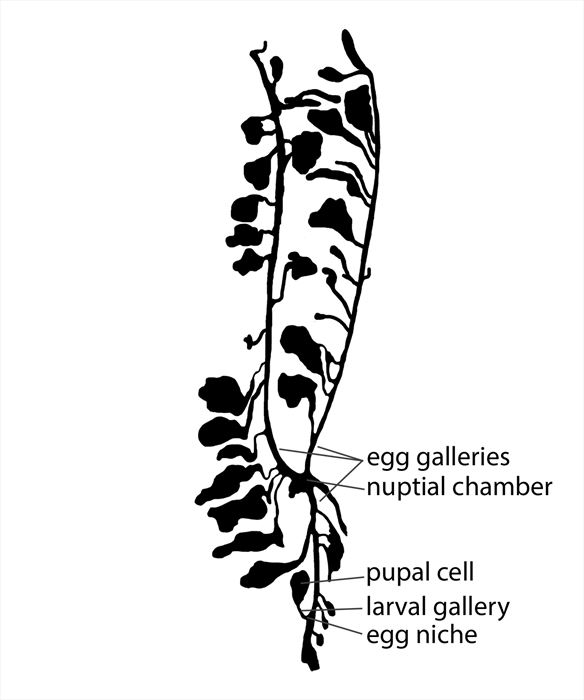
Ips pini, reproductive gallery (under pine bark).
pronotum: The dorsal part of the prothorax. The prothorax is the segment into which the head and first pair of legs are joined.
scutellar sheild: The small exposed dorsal part of the mesothorax between the anterior edges of the elytra and the posterior edge of the pronotum.
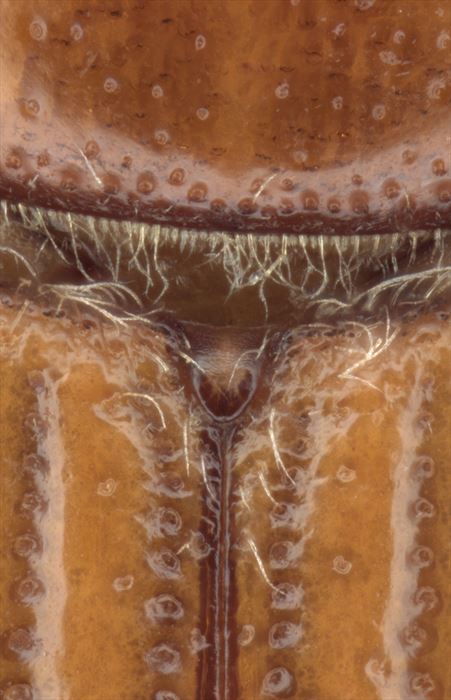
Ips confusus, with scutellar shield at centre between elytra at anterior (top) edge.
socketed teeth:
Spine-like projections in cavities in the protibia. True spines are not separated from the integument of the tibia by any line or groove.
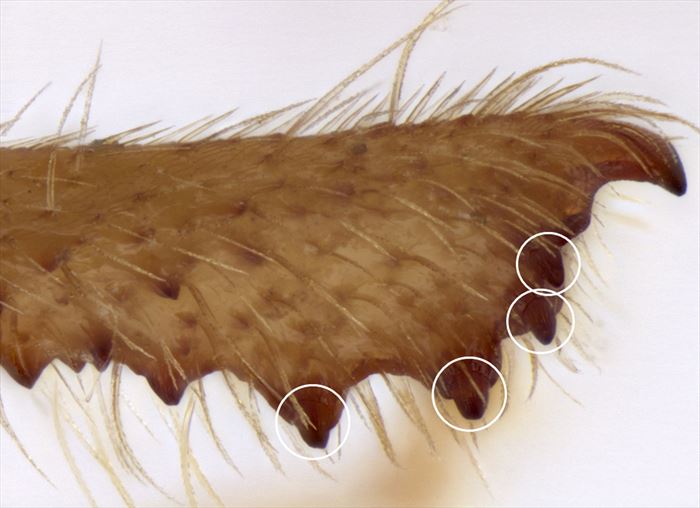
![]()
Biological Survey of Canada
Commission biologique du Canada
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}