
Abstract
The 13 genera of the family Clusiidae (Diptera: Schizophora) are keyed with all characters illustrated and/or photographed. Keys are provided in two forms: a traditional dichotomous key and a non-dichotomous Lucid key. The 36 species of Clusiidae (the six species of the Clusiodes melanostomus complex are treated as a single unit) occurring in Canada and the United States are similarly keyed and illustrated. Genus and species pages are given for each taxon, providing photo galleries, descriptive notes and distributional maps.

Clusia lateralis. Photo by D.K.B. Cheung
General
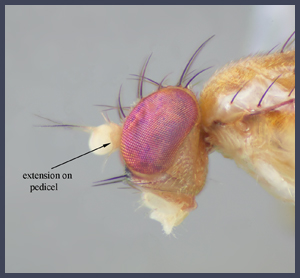
Clusiids are characteristically slender acalyptrate flies usually 2.5-6.0 mm in length, most readily identified by an angulate extension on the outer margin of the pedicel, a dorsoapical arista (dorsobasal in other similar families) on an orbicuar first flagllomere, a complete subcosta, one subcostal break (indistinct in most Clusiodinae), one pair of vibrissae, and five or fewer fronto-orbital bristles. Most species are yellow with a brown to black pattern, but several species are predominantly to entirely pale (some Sobarocephala Czerny), and some are predominantly brown to black (many Heteromeringia Czerny and Craspedochaeta Czerny).
Chaetotaxy
There are 3-5 (sometimes 2) fronto-orbital bristles, of which the anterior is sometimes inclinate, and in species with 4-5 bristles the third from the back is sometimes inclinate and proclinate. Interfrontal bristles are convergent or absent. Ocellar and postvertical bristles are either divergent or absent. On the thorax, there are 1-3 dorsocentral bristles, 1 postpronotal, 2 notopleurals, 2-3 intra-alars (the presutural intra-alar is sometimes absent), 1-2 intrapostalars, and 1 strong anepisternal and katepisternal bristle; 1 pair of prescutellar acrostichal bristles sometimes present. Preapical tibial bristles either absent, present on mid tibia only, or present on mid and hind tibiae.
Habitat
Adults are not often collected, but they can be relatively abundant in some microhabitats. Small dung baits and Malaise traps have been the most successful collection methods , but a number of species have been swept from grass or collected from foliage, logs and dead patches on tree trunks. North American adults appear to prefer mixed and deciduous forests sometimes associated with grass-dominated areas. Tropical species have often been collected along waterways in mossy, humid habitats. Species of Heteromeringia seem to prefer more open forests and have been found in treefalls (Lonsdale & Marshall, 2007a). Clusiids have been known to feed on nectar, rotting vegetative matter, sap (Soós, 1987), and the dung of birds and mammals.

Biology
Clusiidae are among the few acalyptrates that form leks. Males establish dominance at a lek site, defending mating territories (devoid of resources) from other males. True lekking behaviour has only been seen in the subfamily Clusiodinae so far (see photos of Craspedochaeta pullipleura Lonsdale & Marshall from Bolivia), but indirect evidence, such as antlers in Procerosoma Lonsdale & Marshall and widened heads in Sobarocephala, suggest that similar male agonistic behaviour is more widespread in the family.
After mating, the females lay eggs elsewhere, usually under bark or in wood in a state of more advanced decay than the dry surface used for lekking (Roháček, 1995). Selection of clusiid oviposition sites does not appear to be associated with any particular species of tree, but is limited by "humidity, amount of shade, stage of wood decay, [and the] presence of mycelia of certain fungi" (Roháček, 1995). Rearing records suggest that most temperate species can be found in a number of different deciduous trees, but some Clusiodes have also been found in conifers (Roháček, 1995) and larvae of Sobarocephala have been found termite colonies as well as decaying wood (Sóos, 1987). Malloch (1918) found the larvae of S. flaviseta to be "evidently associated with the burrows of coleopterous insects" and to be relatively sluggish and slow moving. Clusiid larvae are probably at least facultatively predaceous on larvae of other wood-boring insects.
As discussed above, males of some clusiids, particularly Hendelia Czerny, have enlarged heads or other conspicuous modifications used in fighting or mutual assessment on lek sites (McAlpine, 1976; Marshall, 2000). Sobarocephala latipennis Melander & Argo and several Australian Hendelia have strongly widened heads, and Hendelia kinetrolicros (Caloren & Marshall), Hendelia mirabilis (Frey), and Procerosoma alini (Shatalkin) have spectacular long genal processes that are probably used in male-male agonistic interactions, although these species have never been observed while engaged in such behaviour. Analogues of the widened head can be seen in the North American species Heteromeringia nitida, which holds its contrastingly coloured fore legs out to the side of the head and folds its tibiae backwards, using these bent legs to exaggerate body size.
General.

Clusiids are characteristically slender acalyptrate flies usually 2.5-6.0 mm in length, most readily identified by an angulate extension on the outer margin of the pedicel, a dorsoapical arista (dorsobasal in similar families) on an orbicuar first flagllomere, a complete subcosta, one subcostal break, one pair of vibrissae, and five or fewer fronto-orbital bristles. Most species are yellow with a brown to black pattern, but several species are entirely pale (some Sobarocephala Czerny), and a number of taxa are predominantly brown to black (many Heteromeringia Czerny and Craspedochaeta Czerny).
Chaetotaxy. There are three to five (sometimes two) fronto-orbital bristles; the anterior bristle is sometimes inclinate, and if there are four or five fronto-orbitals, the third from the back is sometimes inclinate and proclinate (Craspedochaeta). Convergent interfrontal bristles are sometimes present. The ocellar and postvertical bristles are divergent and usually small to absent. On the thorax, there are one to three dorsocentral bristles, one postpronotal, two notopleurals, two or three intra-alars (the presutural intra-alar is sometimes absent), one or two intrapostalar bristles, and one strong anepisternal and katepisternal bristle; a presutural intra-alar and/or a prescutellar acrostichal is sometimes present. The mid and hind tibiae sometimes have one or two dorsal preapical bristles. The subcosta is complete and a subcostal break is present, although this break is indistinct in most Clusiodinae.

 ♂ Genitalia. Externally, clusiid male genitalia are composed of a dome-shaped epandrium, well developed surstyli, two cerci that are confluent basally, and an 'annulus' comprised of sternites 6-8. Internally, the genitalia are made up of a subepandrial sclerite, a hypandrium with three lateral setae and a pair of 'arms' that attach to the subepandrial sclerite, a rod-like phallapodeme, a ring-like basiphallus, a fin-like epiphallus, a short to long distiphallus, one pair of lateral lobes at the base of the distiphallus, a single ejaculatory apodeme, and one pair of pregonites and postgonites. This ground-plan is highly modified in the Clusiodinae excluding Allometopon Kertesz, as the postgonite, epiphallus, and lateral lobes of the distiphallus are absent, the pregonite is enlarged and fused to the hypandrium, the phallapodeme is short and variably modified, and the hypandrial arms are partly attached to the annulus (Lonsdale & Marshall, 2006). The phallus itself may also be highly modified, being either large and sac-like, extremely long and coiled, or atrophied to absent.
♂ Genitalia. Externally, clusiid male genitalia are composed of a dome-shaped epandrium, well developed surstyli, two cerci that are confluent basally, and an 'annulus' comprised of sternites 6-8. Internally, the genitalia are made up of a subepandrial sclerite, a hypandrium with three lateral setae and a pair of 'arms' that attach to the subepandrial sclerite, a rod-like phallapodeme, a ring-like basiphallus, a fin-like epiphallus, a short to long distiphallus, one pair of lateral lobes at the base of the distiphallus, a single ejaculatory apodeme, and one pair of pregonites and postgonites. This ground-plan is highly modified in the Clusiodinae excluding Allometopon Kertesz, as the postgonite, epiphallus, and lateral lobes of the distiphallus are absent, the pregonite is enlarged and fused to the hypandrium, the phallapodeme is short and variably modified, and the hypandrial arms are partly attached to the annulus (Lonsdale & Marshall, 2006). The phallus itself may also be highly modified, being either large and sac-like, extremely long and coiled, or atrophied to absent.
 ♀ Genitalia. The female genitalia are much simpler than those of the male, being comosed of a ventral receptacle and one pair of spermathecae. These structures are weakly sclerotized and difficult to examine, and staining is often necessary. In the Clusiinae and Sobarocephalinae, the spermathecae are often simple and spherical, but in the Clusiodinae, they are longitudinally segmented and sometimes telescoped and heavily pigmented. The ventral receptacle is usually sac-like and recurved, but this generalized form is variably modified in several genera.
♀ Genitalia. The female genitalia are much simpler than those of the male, being comosed of a ventral receptacle and one pair of spermathecae. These structures are weakly sclerotized and difficult to examine, and staining is often necessary. In the Clusiinae and Sobarocephalinae, the spermathecae are often simple and spherical, but in the Clusiodinae, they are longitudinally segmented and sometimes telescoped and heavily pigmented. The ventral receptacle is usually sac-like and recurved, but this generalized form is variably modified in several genera.
Eggs. Eggs are translucent, usually three to four times longer than wide, and approximately as long as sternite 6 of the female (Lonsdale & Marshall, in press). Both ends are tapered (most pronounced anteriorly) and the micropyle is small and terminal. The surface of the egg is usually minutely tuberculate with (usually) no more than a dozen longitudinal wrinkles, but there are numerous departures from this generalized state. In Sobarocephala, these wrinkles are bordered by larger quadrate tubercles, and some of the wrinkles are branched.
Larvae. Larvae are known from European Clusiodes Coquillett. The cephalic papillae are minute, the mandibles are small and well sclerotized, and the cephalopharyngeal skeleton is vestigial and unpigmented (Soós, 1987). The anterior spiracles have six openings, each on a small raised tubercle (Soós, 1987). The posterior spiracles are elevated on chitinized plates, produced as dorsally curving hooks with three elongate-oval openings on the medial or ventral half (Malloch, 1918). The anal plate is large, heavily sclerotized, wider than long and tapered laterally (Malloch, 1918).
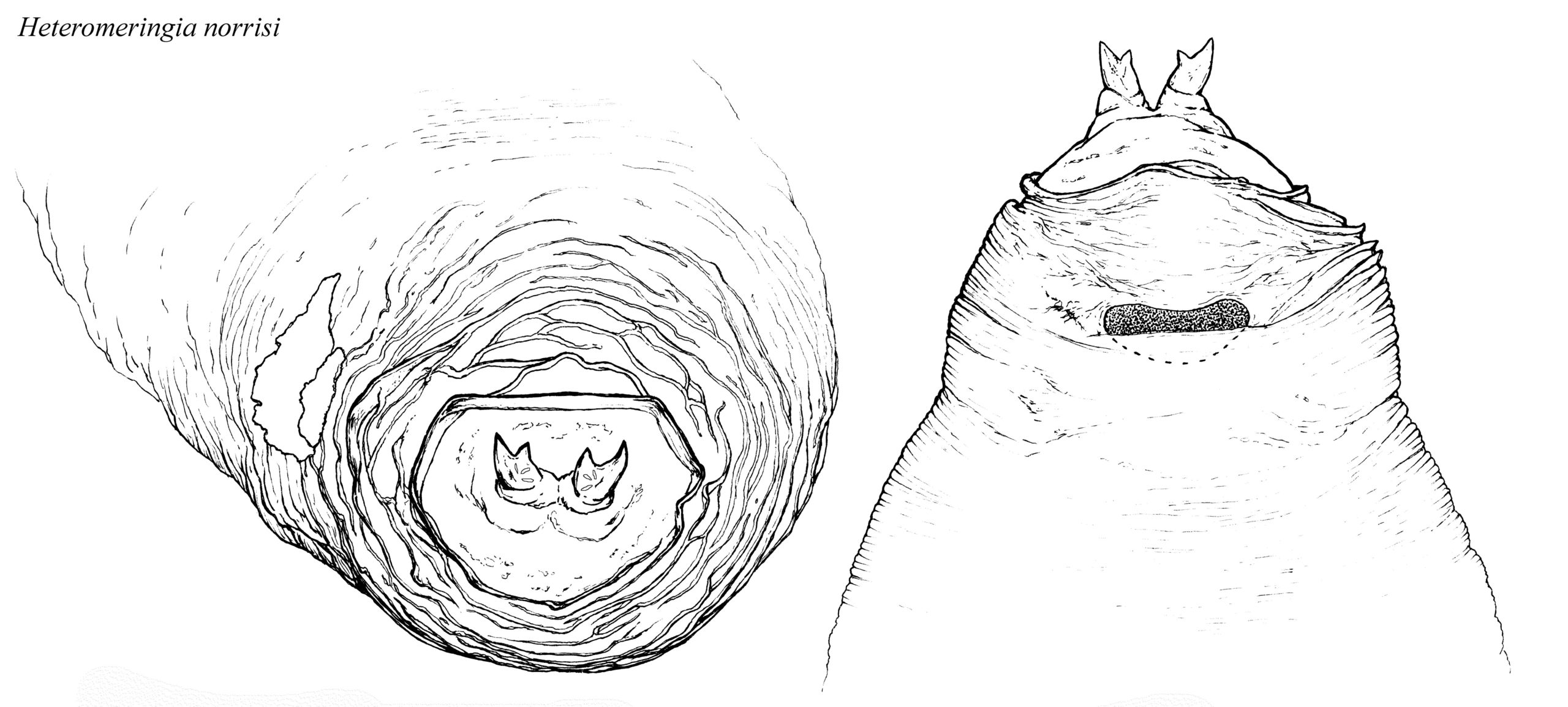
Puparia. Puparia (known from temperate Clusia Haliday, Sobarocephala and Clusiodes) are brownish-orange in colour and covered with numerous minute transverse wrinkles (Lonsdale & Marshall, in press). The posterior face is smooth with various sculpturing, and either sharply curved and bordered by a high ridge (Clusia and Sobarocephala), or broadly rounded with the ridge broken and shallow (Clusiodes). The anal hooks curve dorsally (Clusiodes) or dorsomedially, and are ovate (Clusia) or semi-circular (Sobarocephala) in cross-section. The puparium of the Australian Heteromeringia norrisi is unusual in having an additional inner triangular process on the posterior spiracles.

Eggs, Heteromeringia spp.

Figure: Immature stages clockwise from left: egg, Clusiodes pictipes (Zetterstedt); puparium (posterior face), Clusiodes johnsoni Malloch; puparium (posterior face), Clusiodes melanostomus (Loew); puparium (posterior face), Clusia occidentalis Malloch; puparium (dorsal - posterior third), Clusia occidentalis; puparium (lateral) Clusiodes johnsoni.
 Unless otherwise indicated on the photographs, all photographs of living specimens were taken by SM. We thank the following other photographers for permission to use their work (copyright remains with the photographer): Dmitry Gavryushin (Clusia tigrina, Hendelia beckeri), Tom Murray (Clusiodes melanostomus complex) and John Klymko (lekking Clusiodes). Geoff Thompson photographed the holotype of Allometopon perkinsi and Chris Ho provided the photographs used for the habitus plate of Heteromeringia czernyi. DC took the dorsal photograph of Melanoclusia bivibrissa. Genitalic illustrations of Clusiodes species, excluding the labeled figure provided by OL, were produced by David Caloren. All remaining illustrations and photographs of preserved specimens were taken by OL.
Unless otherwise indicated on the photographs, all photographs of living specimens were taken by SM. We thank the following other photographers for permission to use their work (copyright remains with the photographer): Dmitry Gavryushin (Clusia tigrina, Hendelia beckeri), Tom Murray (Clusiodes melanostomus complex) and John Klymko (lekking Clusiodes). Geoff Thompson photographed the holotype of Allometopon perkinsi and Chris Ho provided the photographs used for the habitus plate of Heteromeringia czernyi. DC took the dorsal photograph of Melanoclusia bivibrissa. Genitalic illustrations of Clusiodes species, excluding the labeled figure provided by OL, were produced by David Caloren. All remaining illustrations and photographs of preserved specimens were taken by OL.
We thank the editorial staff at the journals Studia Dipterologica, Zootaxa, Beiträge zur Entomologie, Bishop Museum Occasional Papers and the European of Journal Entomology for permission to reprint select illustrations from articles previously published by OL and SM.
Funding for this project was provided by an NSERC Discovery grant awarded to SM, an NSERC grant awarded to OL, and support from the Biological Survey of Canada Foundation for DC's role as technical editor of CJAI.
Barraclough, D.A. 2000. The identity of Strongylophthalmyia Heller species (Diptera: Schizophora: Strongylophthalmyiidae) described from the Afrotropical Region, and their transfer to the family Clusiidae. Annals of the Natal Museum 41: 103-106.
Barraclough, D.A. 2002. A new species of Tetrameringia McAlpine (Diptera: Schizophora: Clusiidae) from Malawi, the third species from the Afrotropical Region. African Invertebrates 43: 5-10.
Caloren D.C. & Marshall, S.A. 1998. A revision of the New World species of Clusiodes Coquillett (Diptera: Clusiidae). Studia Dipterologica 5: 261-321.
Frey, R. 1960. Studien über indoaustralische Clusiiden (Dipt.) nebst Katalog der Clusiiden. Commentationes Biologicae 22(2): 1-31.
Greve, L. & Midtgaard, F. 1986. The Clusiidae (Diptera) from the islands Håøya and Ostøya in the Oslofjord and a survey of the family in Norway. Fauna Norvegica (Series B) 33: 86-92.
Lonsdale, O. & Marshall, S.A. 2006a. Redefinition of the Clusiinae and Clusiodinae, description of the new subfamily Sobarocephalinae, revision of the genus Chaetoclusia and a description of Procerosoma gen. n. (Diptera: Clusiidae). European Journal of Entomology 103: 163-182.
Lonsdale, O. & Marshall, S.A. 2006b. Revision of the New World Craspedochaeta Czerny. Zootaxa 1291: 1-101.
Lonsdale, O. & Marshall, S.A. 2007a. Revision of the New World Heteromeringia (Diptera: Clusiidae: Clusiodinae). Beiträge zur Entomologie 57(1): 37-80.
Lonsdale, O. & Marshall, S.A. 2007b. Redefinition of the genera Clusiodes and Hendelia (Diptera: Clusiidae: Clusiodinae), with a review of Clusiodes. Studia Dipterologica 14: 117-159.
Lonsdale, O. & Marshall, S.A. 2007c. Revision of the North American Sobarocephala (Diptera: Clusiidae, Sobarocephalinae). Journal of the Entomological Society of Ontario 138: 65-106.
Lonsdale, O. & Marshall, S.A. 2007d. Revision of the genus Phylloclusia (Diptera: Clusiidae: Clusiinae). Canadian Entomologist 138: 778-792.
Lonsdale, O. & Marshall, S.A. 2008a. Synonymy within Clusia and description of the new genus Melanoclusia (Diptera: Clusiidae: Clusiinae). Annals of the Entomological Society of America 101(2): 327-330.
Lonsdale, O. & Marshall, S.A. 2008b. The Clusiidae (Diptera: Schizophora) of Fiji, with redefinition of Heteromeringia Czerny and synonymy of Tranomeringia Sasakawa. In: Evenhuis, N.L. & Bickel, D.J. (eds.), Fiji Arthropods XI. Bishop Museum Occasional Papers 98: 3-14.
Lonsdale, O. & Marshall, S.A. 2008c. Revision of the temperate South American genus Apiochaeta Czerny, 1903, with synonymy of Alloclusia Hendel, 1917 (Diptera: Clusiidae). Zootaxa 1944: 1-33.
Malloch J.R. 1918. A revision of the dipterous family Clusiodidae (Heteroneuridae). Proceedings of the Entomological Society of Washington 20(1): 2-8.
Marshall, S.A. 2000. Agonistic behaviour and generic synonymy in Australian Clusiidae (Diptera). Studia Dipterologica 7: 3-9.
McAlpine, D.K. 1960. A review of the Australian species of Clusiidae (Diptera: Acalyptrata). Records of the Australian Museum 25: 63-94.
McAlpine, D.K. 1976. Spiral vibrissae in some clusiid flies (Diptera: Schizophora). Australian Entomological Magazine 3(4): 75- 78.
Melander A.L. & Argo N.G. 1924. Revision of the two-winged flies of the family Clusiidae. Proceedings of the United States National Museum 64: 1-54.
Perry, I. & Stubbs, A.E. 1978. Dead wood and Sap runs. A Dipterist's Handbook 15: 65-73.
Roháček, J. 1995. Clusiidae (Diptera) of the Czech and Slovak Republics: Faunistics and notes on biology and behaviour. Časopis Slezského zemského Muzea (A) 44: 123-140.
Roháček, J. 2005. Clusiidae (různatkoviti). Pp. 326-327. In J. Farkač, D. Král & M. Škorpík (eds), Červený seznam ohrožených druhů České republiky. Bezobratlí. List of threatened species in the Czech Republic. Invertebrates. – Agentura ochrany přírody a krajiny ČR, Praha, 760pp.
Sabrosky, C.W. & Steyskal, G.C. 1974. The genus Sobarocephala (Diptera: Clusiidae) in America North of Mexico. Annals of the Entomological Society of America 67(3): 371-385.
Sasakawa, M. 1971. Studies on the Oriental and Pacific Clusiidae (Diptera), pt. 2. Scientific Reports of the Kyoto Prefectural University, Agriculture 23: 50-63.
Sasakawa, M. 1998. Family Clusiidae. Pp. 219-225. In L. Papp & B. Darvas (eds), Contributions to a Manual of Palaearctic Diptera; 3: Higher Brachycera. Science Herald, Budapest. 880pp.
Smith, K.G.V. 1950. The puparium of Clusia flava Mg. (Diptera: Clusiidae). The Entomologist’s Monthly Magazine 86: 53.
Soós, A. 1981. 57: család: Clusiidae – Fatönlegyek. Fauna Hungariae 15(5): 93-108. [In Hungarian]
Soós, A. 1987. Clusiidae. Pp. 853-857. In McAlpine J.F. (ed.), Manual of Nearctic Diptera, Volume 2. Monograph 28, Research branch, Agriculture Canada, Ottawa. 1332pp.
Stubbs, A.E. 1982. An identification guide to British Clusiidae. Proceedings of the Transactions of the British Entomological Natural History Society 15: 89-93.
Stuckenberg, B.R. 1973. New and little-known Clusiidae (Diptera) from South Africa and Mocambique. Annals of the Natal Museum 21(3): 579-593.
Sueyoshi, M. 2006. Species diversity of Japanese Clusiidae (Diptera: Acalyptrata) with description of 12 new species. Annales de la Société Entomologique de France 42(1): 1-26.
Tuomikoski, R. 1936. Bemerkungen über die Clusiiden (Dipt.) Finnlands. Annals Entomologici fennici 2(4): 182-186.
Withers, P. 1985. Notes on some British Clusiidae and reduction of Clusiodes facialis (Coll.) to synonymy. Proceedings of the Transactions of the British Entomological Natural History Society 18: 63-64.
![]()
Biological Survey of Canada
Commission biologique du Canada
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}