
Ambush bugs of the genus Phymata Latreille, 1802 (Heteroptera: Reduviidae: Phymatinae) are some of the most recognizable assassin bugs found in North America. Despite their charismatic morphology and ubiquity, the taxonomy of the Nearctic fauna has not been reviewed in more than half a century. Many species are challenging to identify due to their striking intraspecific variation and sexual dimorphism and only subtle interspecific differences. Considering this difficult taxonomy and that recent molecular phylogenetic studies have demonstrated para- or polyphyly among currently recognized species-level taxa, reclassification is required. Ambush bugs of the Nearctic erosa species group are herein revised. Eleven previously recognized species are redescribed: Phymata americana Melin, 1931; Phymata arctostaphylae Van Duzee, 1914; Phymata borica Evans, 1931; Phymata fasciata (Gray, 1832); Phymata granulosa Handlirsch, 1897; Phymata luteomarginata Kormilev, 1957; Phymata pacifica Evans, 1931; Phymata pennsylvanica Handlirsch, 1897; Phymata rossi Evans, 1931; Phymata saileri Kormilev, 1957; and Phymata salicis Cockerell, 1900. Five subspecies are elevated to species rank and redescribed: Phymata mystica Evans, 1931 stat. restit.; Phymata metcalfi Evans, 1931 stat. restit.; Phymata mexicana Melin, 1931 stat. nov.; Phymata obscura Kormilev, 1957 stat. nov.; and Phymata stanfordi Evans, 1931 stat. nov. One new species from southern California is described: Phymata paraborica sp. nov. Phymata americana coloradensis Melin, 1931 syn. nov., Phymata pacifica hainesi Kormilev, 1962 syn. nov., Phymata granulosa chiriquiensis Melin, 1931 syn. nov., and Phymata granulosa evansi Kormilev, 1962 syn nov. are each synonymized with their respective nominate subspecies. Phymata fasciata panamensis Kormilev, 1962 syn. nov. is synonymized with Phymata mexicana Melin, 1931. Phymata granulosa texasana Kormilev, 1957 syn. nov., is synonymized with Phymata rossi. An illustrated key to ambush bugs of the United States and Canada, detailed diagnoses, habitus images, and distribution maps are provided for all 17 Nearctic erosa species group taxa. Furthermore, we incorporate citizen science observations of ambush bugs from an iNaturalist.org project with traditional specimen-based data to expand our knowledge of species-level variation and distribution.

Phymata pennsylvanica Handlirsch, 1897 (photo by Nevin Cullen)
The genus Phymata Latreille, 1802 (Heteroptera: Reduviidae) comprises ~110 species of ambush bugs distributed primarily in the New World (Froeschner and Kormilev 1989). Their peculiar morphology, sexual dimorphism, and predation on other flower-associated arthropods have made them popular subjects for evolutionary (Punzalan et al. 2008a; Punzalan et al. 2008b, c, 2010, Weirauch et al. 2011; Punzalan and Rowe 2015, 2016), ecological (Balduf 1941; Mason 1977, 1986; Yong 2005; Masonick et al. 2019), and behavioral research (Dodson and Marshall 1984; Elliott and Elliott 1991, 1994; Yong 2003; Dixon and Rasmussen 2013). While these sit-and-wait predators are easily distinguished from other true bugs by their raptorial forelegs, fusiform antennae, deep antennal excavations of the head and propleura, and dorsolaterally flared pronotum and abdomen, species-level identification within the genus is confounded by subtle interspecific differences, sexual dimorphism, and intraspecific variation. This paradox holds especially true for the Nearctic erosa species group, a clade represented by the predominant ambush bug fauna of North America. While the vast majority of Phymata found in the wild and in collections belong to just a few common and widespread erosa group species, many rare endemic species are found in the deserts and scrublands of the western United States and northern Mexico.
The Nearctic erosa group, and Phymata as whole, have a convoluted taxonomic history. Taxonomic research on ambush bugs began when Linnaeus in 1758 described the first species, Cimex erosus, based on a specimen from Surinam. Latreille erected the genus Phymata in 1802 and reclassified C. erosus as Phymata erosa in 1804, a name under which numerous Neotropical and Nearctic subspecies would later be classified. Handlirsch (1897) provided the first monograph to focus exclusively on Phymatinae, and subsequent revisions by Melin (1931), Evans (1931), and Kormilev (1962) further refined understanding of their diversity, and over time, all Nearctic P. erosa subspecies were elevated to species rank. In addition to biological issues such as polymorphic traits and sexual dimorphism that hinder species diagnosis, the taxonomy of the erosa group has been complicated by the loss or lack of designated types, vague and/or poorly illustrated descriptions, and descriptions based on singleton specimens or on only one sex. What is more, coloration, a feature that can be quite variable within populations and one that changes drastically over time both in live and pinned specimens (Schuh and Slater 1995; Boyle and Start 2020), was used in many cases to define taxa. Outdated descriptions and keys, the most recent having been published in 1962, have culminated in numerous specimens in natural history collections being mis- or unidentified. While little has been done to modernize ambush bug taxonomy, molecular phylogenetics and integrative species delimitation have recently shed light on their species-level relationships and revealed that several erosa group taxa are para- or polyphyletic (Masonick et al. 2017; Masonick and Weirauch 2020). Taxonomic updates are sorely needed and insights from these analyses have helped structure the present revision.
The primary aim of this revision is to alleviate confusion pertaining to erosa group taxa by providing illustrated keys and detailed species diagnoses. To circumvent the issue of subtle or polymorphic characters and increase the chances of correct identification, our diagnoses reference suites of traits relevant for identifying each taxon, and do not rely on any single attribute. Prior to this project, 11 erosa group species and 11 subspecies were known from the Nearctic (Kormilev 1962; Froeschner and Kormilev 1989). Herein, we recognize 17 Nearctic erosa species partly through the synonymization or elevation of subspecies to full species of the taxa cataloged by Froeschner and Kormilev. Eleven previously accepted species are redescribed, five subspecies are elevated to species rank and redescribed, and one new species from southern California is described: Phymata paraborica sp. nov. Phymata americana coloradensis Melin, 1931 syn. nov., Phymata pacifica hainesi Kormilev, 1962 syn. nov., Phymata granulosa chiriquiensis Melin, 1931 syn. nov., and Phymata granulosa evansi Kormilev, 1962 syn. nov. are each synonymized with their respective nominate subspecies. Phymata fasciata panamensis Kormilev, 1962 syn. nov. is synonymized with Phymata mexicana Melin, 1931. Phymata granulosa texasana Kormilev, 1957 syn. nov. is synonymized with Phymata rossi Evans, 1931.
Provided in this revision are two illustrated keys to Nearctic ambush bugs. The first (Key(1)), covers rare non-erosa group Phymata native to the United States and Canada. The second (Key(2)) deals exclusively with the 17 species of the Nearctic erosa group clade. Detailed diagnoses, habitus plates, and distribution maps for these taxa are also included. Our treatment of the Nearctic erosa group benefited from the examination of more than 4,125 specimens from 14 natural history collections and survey of 2,290 (as of October 2019) transcontinental citizen science observations from our iNaturalist project “Uncovering the ambush bugs” (https://www.inaturalist.org/projects/uncovering-the-ambush-bugs).
For a list of all Phymata known from Canada, the United States, and/or Mexico, see Table 1. Ambush bugs native to North America that are not part of the erosa group clade are excluded from this revision. It should be noted that Phymata severini Handlirsch, 1897 and Phymata parva Handlirsch, 1897 are two taxa from Mexico that were originally erected as subspecies of P. erosa and later elevated to species rank by Melin (1931) and Kormilev (1962), respectively. Based on molecular analysis, we found these two species to both be distantly related to the clade of Nearctic ambush bugs treated here (see Masonick and Weirauch, 2020, Figs S3–S6).
Table 1. Checklist of Phymata found in Canada, the United States, and/or Mexico.
| Phymata Latreille, 1802 (Heteroptera: Reduviidae) |
| Phymata albopicta Handlirsch, 1897 (USA, MEX) |
| Phymata annulipes Stål, 1862 (MEX) |
| Phymata barberi Kormilev, 1962 (MEX) |
| Phymata brailovskyi Kormilev, 1990 (MEX) |
| Phymata luxa Evans, 1931 (USA, MEX) |
| Phymata maculata Kormilev, 1957 (USA) |
| Phymata noualhieri Handlirsch, 1897 (USA, MEX) |
| Phymata pallida Kormilev, 1957 (USA) |
| Phymata parva Handlirsch, 1897 (MEX) |
| Phymata rhynocerata Kormilev, 1957 (MEX) |
| Phymata severini Handlirsch, 1897 (MEX) |
| Phymata turnbowi Kormilev, 1983 (MEX) |
| Phymata vicina Handlirsch, 1897 (CAN, USA) |
| erosa species group clade |
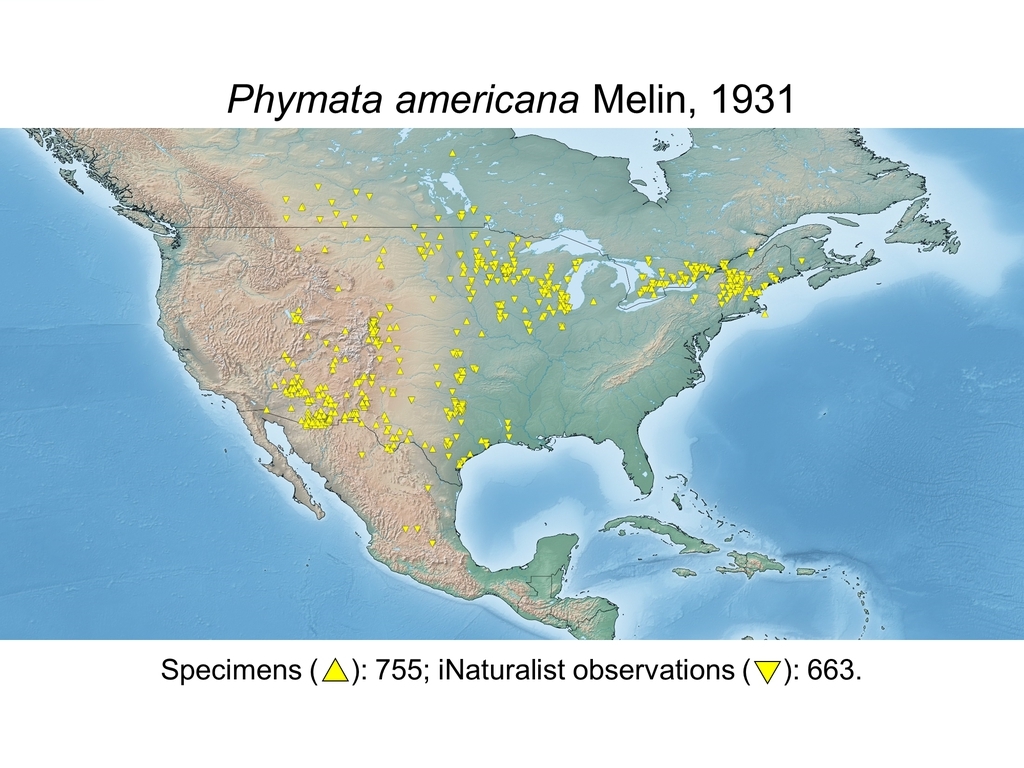
| Phymata americana Melin, 1931 (CAN, USA, MEX) |
| Phymata arctostaphylae Van Duzee, 1914 (USA, MEX) |
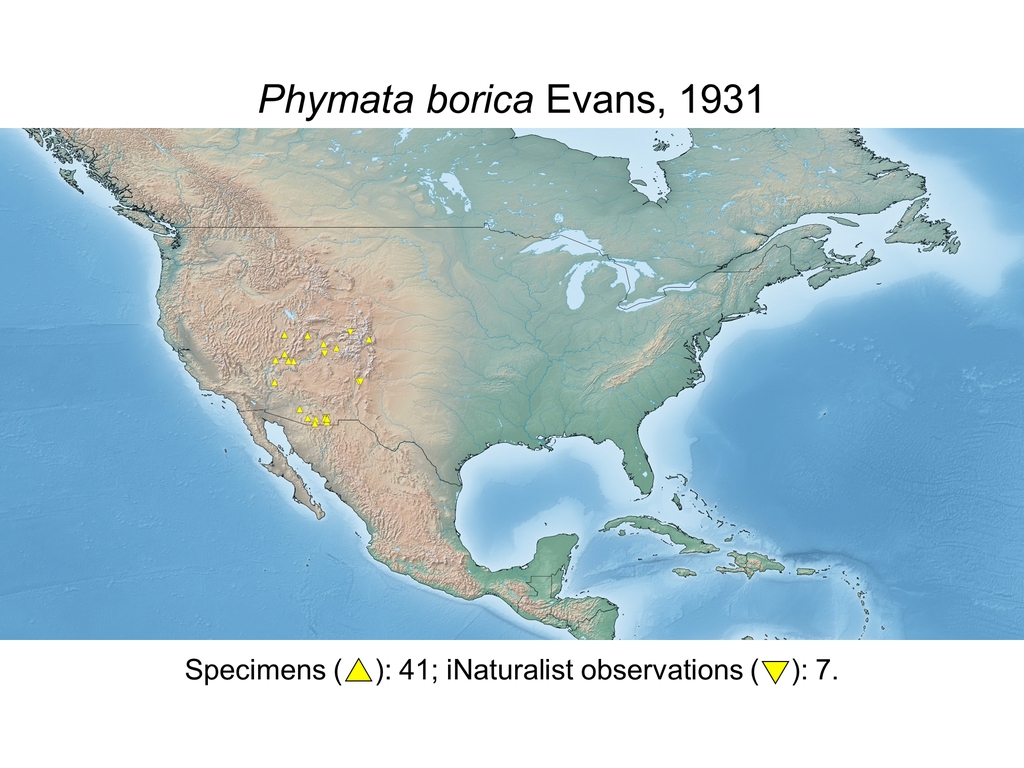
| Phymata borica Evans, 1931 (USA) |
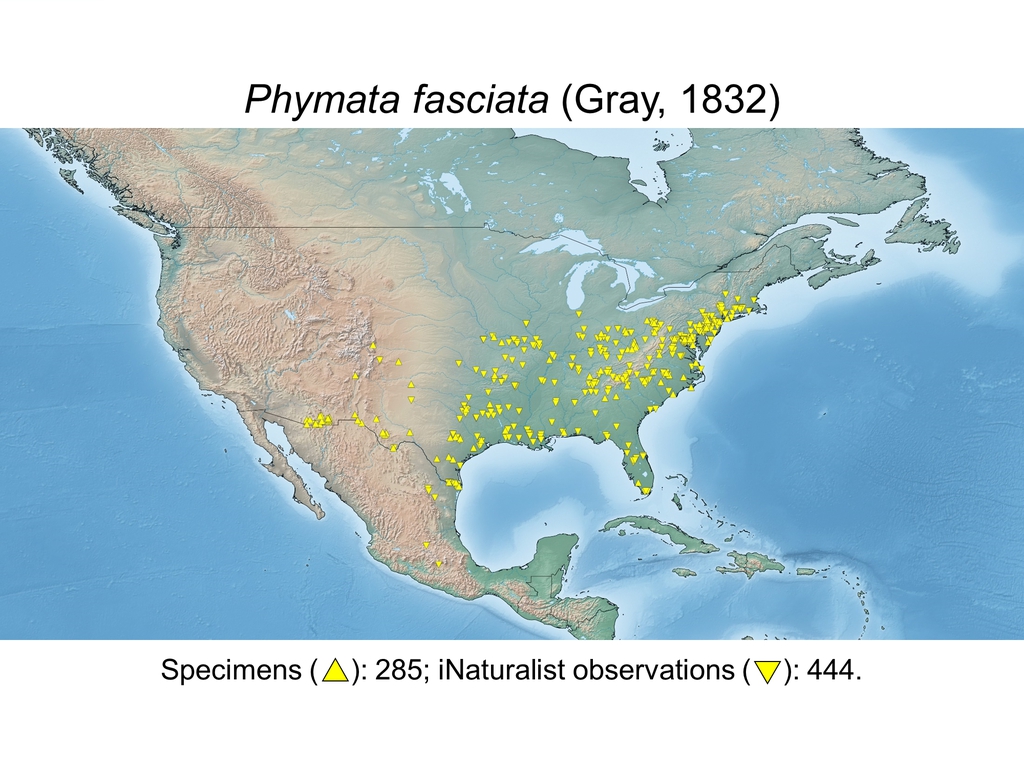
| Phymata fasciata (Gray, 1832) (USA, MEX) |
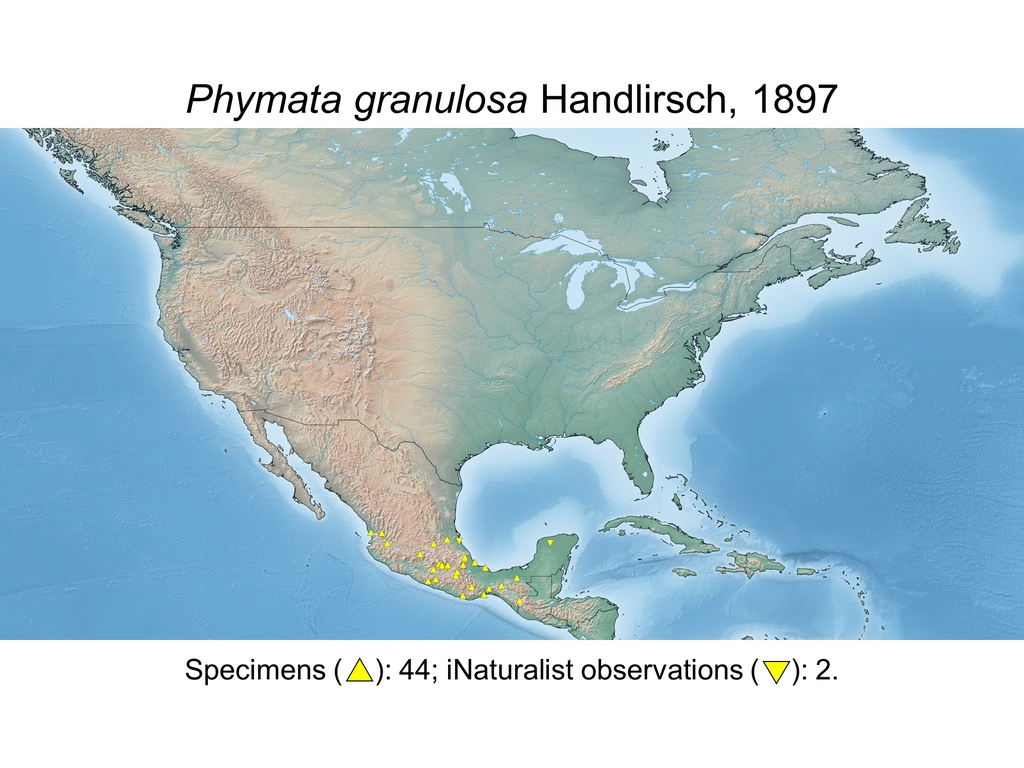
| Phymata granulosa Handlirsch, 1897 (MEX) |
| Phymata luteomarginata Kormilev, 1957 (USA) |
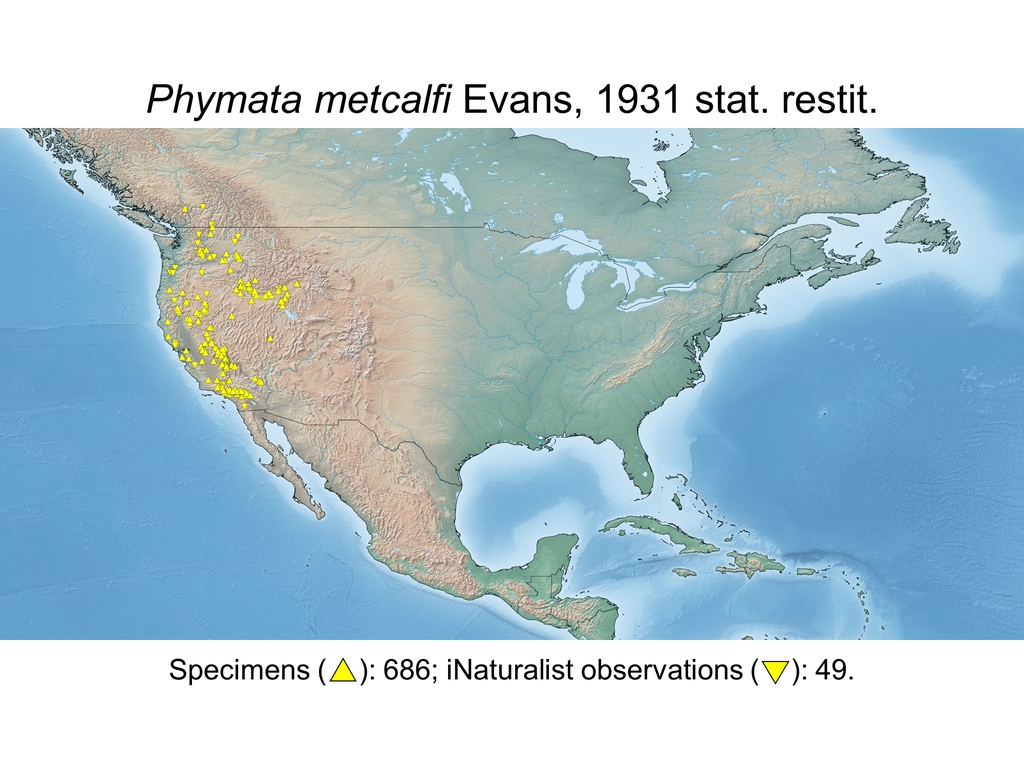
| Phymata metcalfi Evans, 1931 stat. restit. (CAN, USA) |
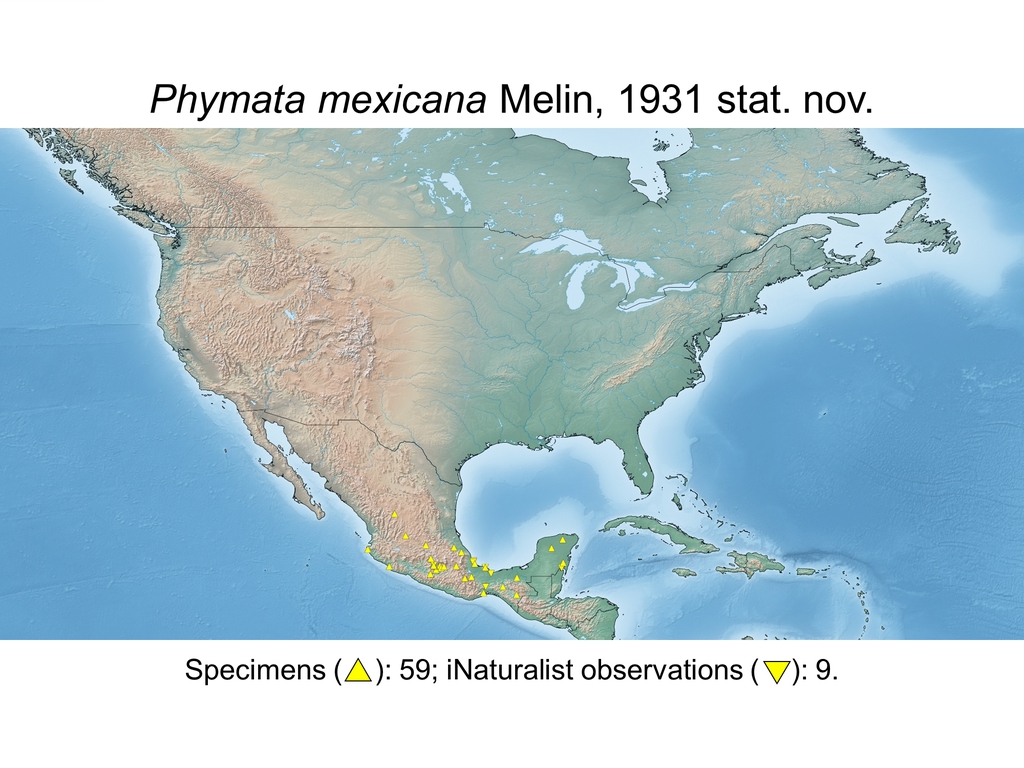
| Phymata mexicana Melin, 1931 stat. nov. (MEX) |
| Phymata mystica Evans, 1931 stat. restit. (USA) |
| Phymata obscura Kormilev, 1957 stat. nov. (USA) |
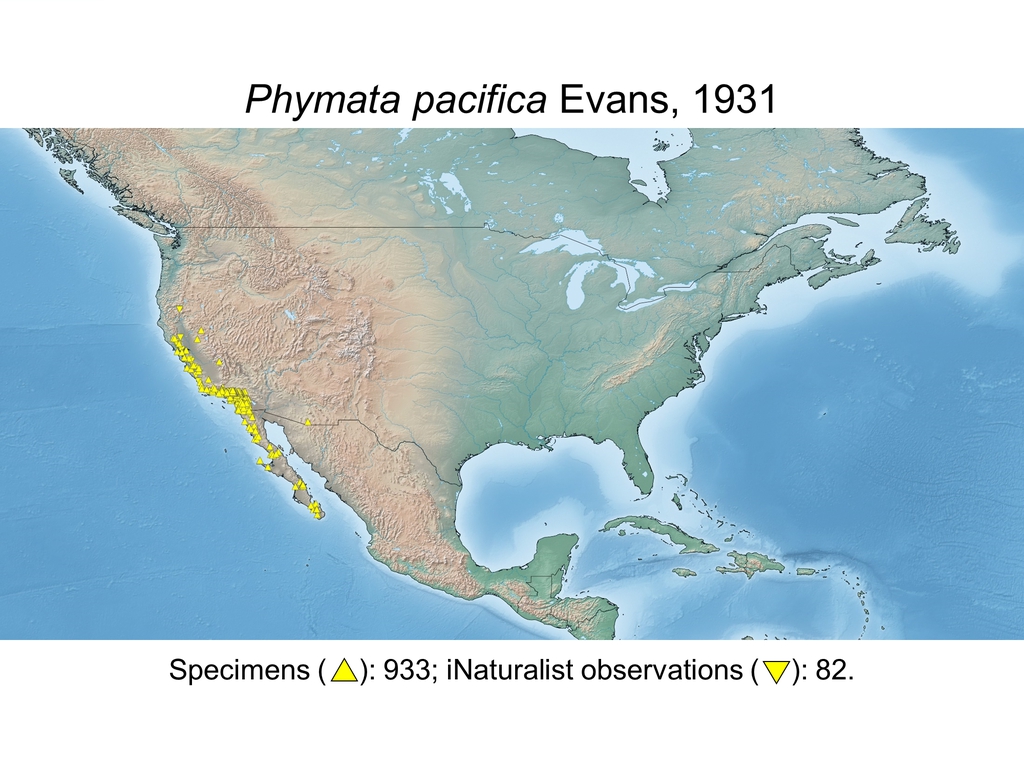
| Phymata pacifica Evans, 1931 (USA, MEX) |
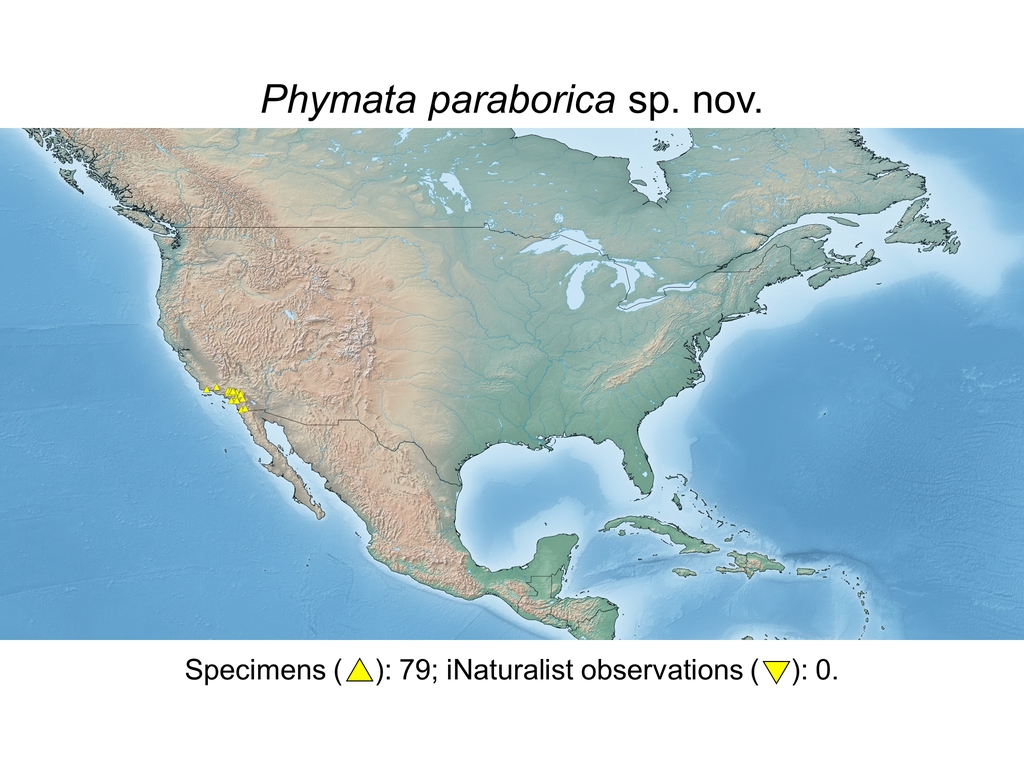
| Phymata paraborica sp. nov. (USA) |
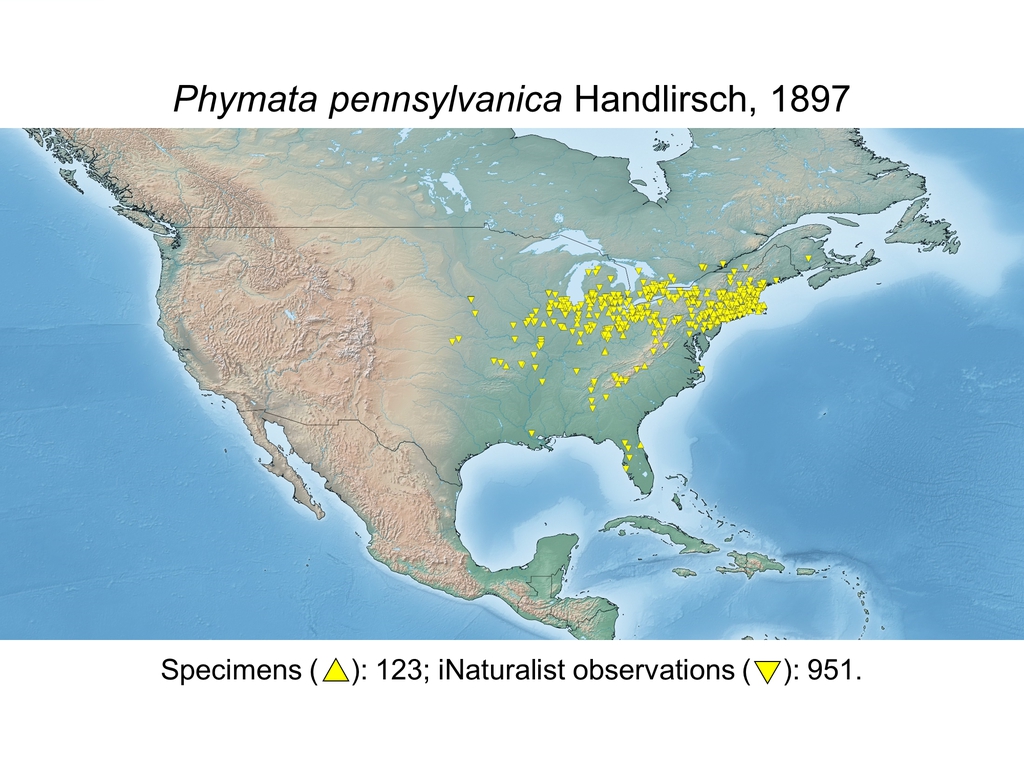
| Phymata pennsylvanica Handlirsch, 1897 (CAN, USA) |
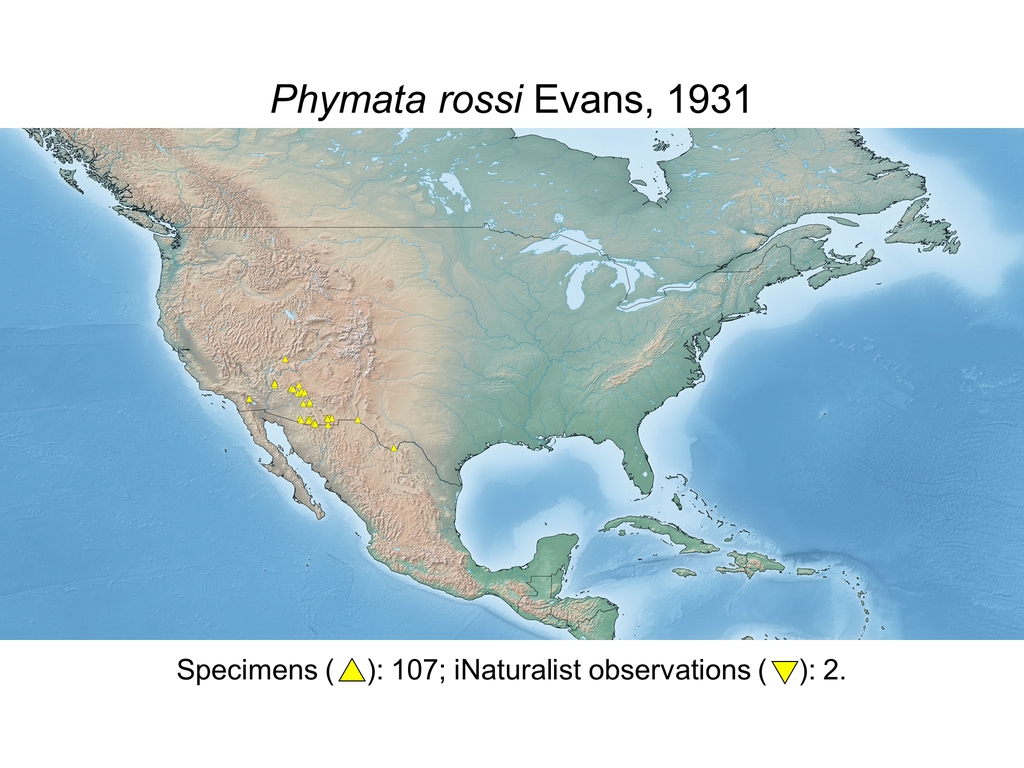
| Phymata rossi Evans, 1931 (USA) |
| Phymata saileri Kormilev, 1957 (USA) |
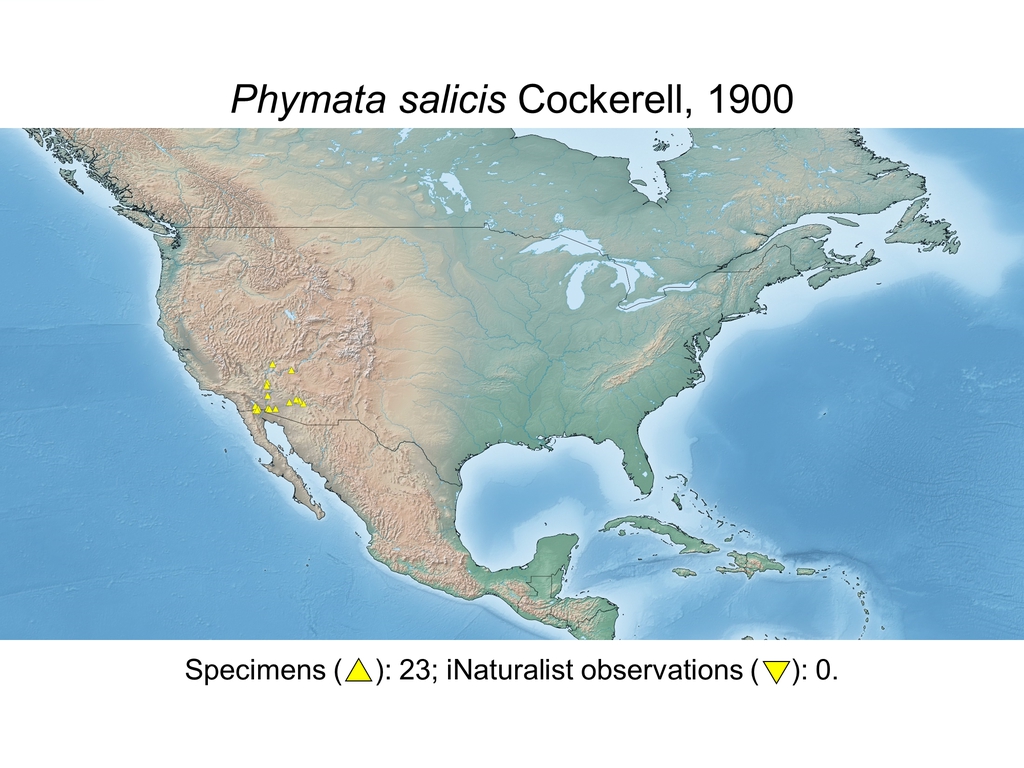
| Phymata salicis Cockerell, 1900 (USA) |
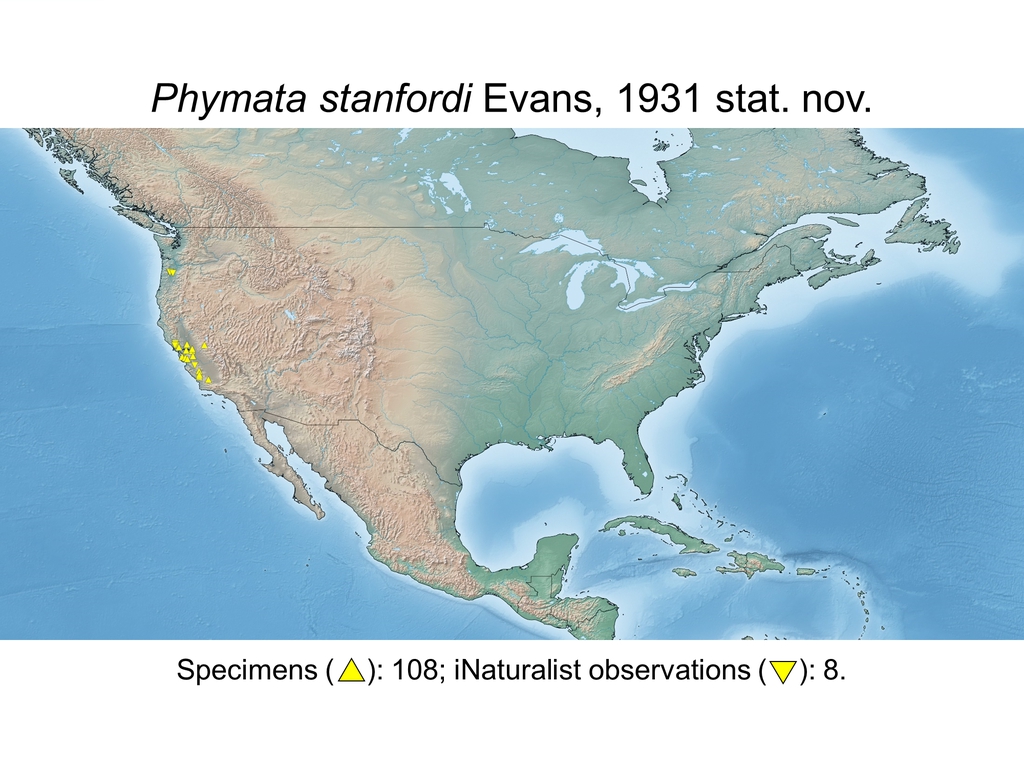
| Phymata stanfordi Evans, 1931 stat. nov. (USA) |
Specimens, depositories, and databasing: A total of 4,125 databased erosa group specimens were examined, of which 1,694 were adult females and 2,385 adult males. Table 2 lists the collections from which material was borrowed and/or is now deposited. Specimens examined are listed by species in the Appendix. We only report material which has been georeferenced: several aberrant individuals which could not be identified or were labeled as being collected drastically out of their known/previously documented range were excluded. We georeferenced older specimens for which geographic coordinates and elevation had not been recorded with locality information using GEOLocate software (https://www.geo-locate.org/) and Google Earth Pro v7.3.2.5776. Specimens were each affixed with a unique specimen identifier (USI) and databased using the Planetary Biodiversity Inventory instance of the Arthropod Easy Capture Database (https://research.amnh.org/pbi/locality/index.php). Specimen information and images can be accessed through the Heteroptera Species Pages (http://research.amnh.org/pbi/heteropteraspeciespage).
Measurements and imaging: Specimen measurements (in mm, Table 3) were taken using a dissecting scope mounted to a digital micrometer positioning system which was connected to a Microcode II® RS-232 digital readout (Boeckeler Instruments®). We recorded measurements of the total body length, the distances across the lateral and posterior pronotal angles (either of which may represent the widest portion of the thorax), and the individual lengths of the pedicel, basiflagellomere, and distiflagellomere (of the left antenna for all specimens in which it was intact). The ratio of the length of the distiflagellomere to that of the pedicel + basiflagellomere is reported for each taxon. This was calculated by taking the mean across ratios for individual specimens of a given sex. While past taxonomists who dealt with this genus often recorded the maximal width of the abdomen, we omitted this trait from study because it is rather contextual and strongly influenced by how the specimen died and whether it had recently fed and/or was carrying mature eggs. Specimen photographs were taken using a Leica Microsystems imaging system (LAS software v4.3.0), stacked with Zerene Stacker v1.04 to create composite images, and then edited in Adobe Photoshop® CC 2017. Figures were prepared in Adobe Illustrator® CC 2017.
Distribution maps and citizen science data: Specimen-associated locality data and observational data (i.e., geocoordinates and images) shared by the general public with our iNaturalist project “Uncovering the ambush bugs” (https://www.inaturalist.org/projects/uncovering-the-ambush-bugs) were used in conjunction to create more complete estimates of current taxon distributions (data last accessed October 9, 2019). This data is provided in a supplementary spreadsheet (XLSX) and image archive (ZIP; 240 MB via Google Drive). Distribution maps (Maps 1-17) were generated with the available coordinate data for each erosa group species in SimpleMappr (https://www.simplemappr.net/).
Taxonomic descriptions: Descriptions were partially produced using a WinClada v1.61 (Nixon 1999) based character matrix and then manually edited to accommodate variability and any unique traits for each species. Outlines displaying the consensus pronotal shape were generated following the geometric morphometric approach outlined in Masonick and Weirauch (2020) and are included for each sex in all taxon plates. The curvature of the pronotal lobes are best viewed from a dorsolateral perspective with the lateral margin placed roughly perpendicular to the optical axis. Pronotal shape variation is described from this position unless otherwise noted. Adult males are described in their entirety, but for females, only sexually dimorphic traits are described. Based on combined molecular data, we recently showed that the erosa group likely comprises three main species complexes, the americana, borica, and fasciata species complexes (see Masonick and Weirauch, 2020, Fig. 3). While these complexes are fairly well-defined using DNA, no morphological synapomorphies are apparent. Due to the lack of diagnostic characters, we only provide a general description for all erosa group taxa and not the individual species complexes themselves. Species descriptions are each accompanied with a plate highlighting their key diagnostic characters. Given their morphological variability and the fact that both age and diet influence color pattern traits in adults (see Punzalan et al. 2008a), we strongly recommend that readers assess multiple characters when making identifications.
Terminology and abbreviations: Anatomical terminology roughly follows that used in Kormilev’s (1962) revision of the genus. Morphological features of Phymata are illustrated in Figures 1, 2, and 20R. The following abbreviations are included in keys, plates, and/or text: aa, anterior angle; af, antennal furrow; apd, anterior pronotal disk; apl, anterior pronotal lobe; bflg, basiflagellomere; cas2–7, connexiva of abdominal segments 2–7; dflg, distiflagellomere; fcx, forecoxa; ffm, forefemur; fp, frontal process; ftb, foretibia; gp, granulation patch; la, lateral angle; lc, longitudinal carina; ln, lateral notch; mspl, mesopleuron; mtpl, metapleuron; op, ocellar process; pa, posterior angle; pd, pedicel; pop, preocellar process; pp, posterior process; ppd, posterior pronotal disk; ppl, posterior pronotal lobe; prpl, propleuron; py, pygophore; sc, scutellum; sppl, shoulder of the posterior pronotal lobe; tb, transverse band of the corium; tlc, tubercles of longitudinal carina.
Botanical taxonomy: Plant names and authors listed in this revision are based on the Integrated Taxonomic Information System database (itis.gov).
Nomenclature: The nomenclatural acts contained within this work have been registered in ZooBank (http://www.zoobank.org), the official registry of the International Commission on Zoological Nomenclature. The LSID (Life Science Identifier) number of the publication is: urn:lsid:zoobank.org:pub:2E2263AA-F7B4-46B6-A26B-97A9F9CE3628.
Table 2. List of specimen depositories and their collection codes.
| Collection | Location | Code |
|---|---|---|
| American Museum of Natural History | New York City, NY USA | AMNH |
| California Academy of Sciences | San Francisco, CA USA | CAS |
| Illinois Natural History Survey Insect Collection | Champaign, IL USA | INHS |
| University of Kansas Snow Entomological Museum | Lawrence, KS USA | KU |
| Natural History Museum of Los Angeles County | Los Angeles, CA USA | LACM |
| Museum of Northern Arizona | Flagstaff, AZ USA | MNA |
| Montana State University Montana Entomology Collection | Bozeman, MT USA | MTEC |
| Natural History Museum Vienna | Vienna, Austria | NHMW |
| Swedish Museum of Natural History | Stockholm, Sweden | NRM |
| San Diego Natural History Museum | San Diego, CA USA | SDNHM |
| University of California Riverside Entomology Research Museum | Riverside, CA USA | UCR |
| Universidad Autonoma de Mexico Instituto de Biologia | Mexico City, Mexico | UNAM |
| United States National Museum of Natural History | District of Colombia, USA | USNM |
| University of Idaho William F. Barr Entomological Museum | Moscow, ID USA | WFBM |
Table 3. Measurements in mm of erosa group taxa. Abbreviations: body, total body length; pd, pedicel; bflg, basiflagellomere; dflg, distiflagellomere; la, lateral angle of pronotum; pa, posterior angle of pronotum.
| Length | Width | ||||||
|---|---|---|---|---|---|---|---|
| Species | body | pd | bflg | dflg | dflg : pd+bflg | la | |
| Phymata americana | |||||||
| males (N=8) | Mean | 8.81 | 0.62 | 0.66 | 1.15 | 0.89 | 3.37 |
| SD | 0.30 | 0.06 | 0.02 | 0.09 | 0.18 | ||
| Range | 1.00 | 0.16 | 0.07 | 0.25 | 0.59 | ||
| Min. | 8.15 | 0.56 | 0.64 | 1.05 | 3.08 | ||
| Max. | 9.15 | 0.72 | 0.71 | 1.30 | 3.67 | ||
| females (N=8) | Mean | 10.06 | 0.65 | 0.70 | 1.03 | 0.76 | 3.74 |
| SD | 0.42 | 0.04 | 0.07 | 0.09 | 0.33 | ||
| Range | 1.14 | 0.11 | 0.22 | 0.23 | 0.93 | ||
| Min. | 9.59 | 0.59 | 0.54 | 0.92 | 3.38 | ||
| Max. | 10.73 | 0.70 | 0.76 | 1.14 | 4.31 | ||
| Phymata arctostaphylae | |||||||
| males (N=5) | Mean | 8.08 | 0.56 | 0.63 | 1.23 | 1.04 | 3.40 |
| SD | 0.41 | 0.03 | 0.01 | 0.05 | 0.23 | ||
| Range | 0.91 | 0.07 | 0.02 | 0.11 | 0.59 | ||
| Min. | 7.70 | 0.52 | 0.62 | 1.18 | 3.06 | ||
| Max. | 8.61 | 0.59 | 0.64 | 1.29 | 3.65 | ||
| females (N=5) | Mean | 9.47 | 0.62 | 0.68 | 1.02 | 0.78 | 3.73 |
| SD | 0.46 | 0.04 | 0.05 | 0.10 | 0.27 | ||
| Range | 1.18 | 0.11 | 0.13 | 0.27 | 0.66 | ||
| Min. | 8.96 | 0.57 | 0.64 | 0.92 | 3.43 | ||
| Max. | 10.14 | 0.68 | 0.76 | 1.20 | 4.09 | ||
| Phymata borica | |||||||
| males (N=4) | Mean | 7.31 | 0.51 | 0.59 | 1.03 | 0.95 | 2.61 |
| SD | 0.38 | 0.04 | 0.06 | 0.09 | 0.04 | ||
| Range | 0.76 | 0.10 | 0.14 | 0.20 | 0.08 | ||
| Min. | 6.93 | 0.46 | 0.52 | 0.91 | 2.56 | ||
| Max. | 7.69 | 0.56 | 0.66 | 1.11 | 2.65 | ||
| females (N=3) | Mean | 8.40 | 0.55 | 0.62 | 0.91 | 0.78 | 2.99 |
| SD | 0.22 | 0.05 | 0.02 | 0.01 | 0.09 | ||
| Range | 0.43 | 0.09 | 0.04 | 0.02 | 0.18 | ||
| Min. | 8.19 | 0.52 | 0.60 | 0.91 | 2.89 | ||
| Max. | 8.62 | 0.61 | 0.64 | 0.92 | 3.07 | ||
| Phymata fasciata | |||||||
| males (N=6) | Mean | 8.21 | 0.56 | 0.65 | 1.08 | 0.89 | 3.14 |
| SD | 0.49 | 0.03 | 0.03 | 0.06 | 0.15 | ||
| Range | 1.46 | 0.07 | 0.09 | 0.14 | 0.43 | ||
| Min. | 7.61 | 0.53 | 0.61 | 1.01 | 2.97 | ||
| Max. | 9.07 | 0.60 | 0.70 | 1.15 | 3.40 | ||
| females (N=4) | Mean | 9.18 | 0.58 | 0.69 | 0.89 | 0.7 | 3.45 |
| SD | 0.47 | 0.08 | 0.05 | 0.13 | 0.22 | ||
| Range | 0.98 | 0.17 | 0.11 | 0.27 | 0.48 | ||
| Min. | 8.62 | 0.52 | 0.65 | 0.74 | 3.25 | ||
| Max. | 9.60 | 0.69 | 0.76 | 1.01 | 3.73 | ||
| Phymata granulosa | |||||||
| males (N=5) | Mean | 8.50 | 0.56 | 0.69 | 1.09 | 0.87 | 3.38 |
| SD | 0.61 | 0.03 | 0.04 | 0.10 | 0.26 | ||
| Range | 1.63 | 0.08 | 0.11 | 0.25 | 0.61 | ||
| Min. | 7.89 | 0.53 | 0.65 | 0.99 | 3.18 | ||
| Max. | 9.52 | 0.60 | 0.76 | 1.24 | 3.78 | ||
| females (N=5) | Mean | 9.61 | 0.66 | 0.79 | 1.06 | 0.74 | 3.90 |
| SD | 0.51 | 0.05 | 0.03 | 0.10 | 0.28 | ||
| Range | 1.22 | 0.14 | 0.08 | 0.23 | 0.71 | ||
| Min. | 8.89 | 0.59 | 0.75 | 0.93 | 3.57 | ||
| Max. | 10.11 | 0.73 | 0.83 | 1.16 | 4.28 | ||
| Phymata luteomarginata | |||||||
| males (N=5) | Mean | 6.99 | 0.51 | 0.49 | 0.96 | 0.96 | 2.71 |
| SD | 0.44 | 0.05 | 0.05 | 0.13 | 0.28 | ||
| Range | 1.15 | 0.12 | 0.12 | 0.32 | 0.72 | ||
| Min. | 6.30 | 0.44 | 0.46 | 0.75 | 2.27 | ||
| Max. | 7.45 | 0.56 | 0.57 | 1.06 | 2.99 | ||
| females (N=5) | Mean | 7.75 | 0.49 | 0.56 | 0.77 | 0.73 | 2.86 |
| SD | 0.82 | 0.06 | 0.06 | 0.09 | 0.25 | ||
| Range | 1.99 | 0.14 | 0.14 | 0.19 | 0.60 | ||
| Min. | 6.99 | 0.42 | 0.51 | 0.66 | 2.63 | ||
| Max. | 8.97 | 0.56 | 0.64 | 0.85 | 3.23 | ||
| Phymata metcalfi | |||||||
| males (N=6) | Mean | 7.73 | 0.54 | 0.58 | 0.96 | 0.86 | 3.01 |
| SD | 0.54 | 0.03 | 0.04 | 0.12 | 0.30 | ||
| Range | 1.34 | 0.09 | 0.10 | 0.30 | 0.75 | ||
| Min. | 7.16 | 0.51 | 0.53 | 0.83 | 2.68 | ||
| Max. | 8.49 | 0.60 | 0.63 | 1.13 | 3.43 | ||
| females (N=6) | Mean | 8.53 | 0.55 | 0.61 | 0.82 | 0.71 | 3.23 |
| SD | 0.68 | 0.05 | 0.04 | 0.07 | 0.29 | ||
| Range | 1.78 | 0.13 | 0.12 | 0.16 | 0.64 | ||
| Min. | 7.47 | 0.51 | 0.53 | 0.74 | 2.79 | ||
| Max. | 9.25 | 0.64 | 0.65 | 0.90 | 3.44 | ||
| Phymata mexicana | |||||||
| males (N=5) | Mean | 8.43 | 0.62 | 0.66 | 1.14 | 0.89 | 3.55 |
| SD | 0.35 | 0.02 | 0.04 | 0.08 | 0.29 | ||
| Range | 0.88 | 0.05 | 0.12 | 0.18 | 0.77 | ||
| Min. | 8.14 | 0.59 | 0.61 | 1.05 | 3.29 | ||
| Max. | 9.03 | 0.64 | 0.73 | 1.24 | 4.06 | ||
| females (N=5) | Mean | 9.40 | 0.64 | 0.73 | 1.00 | 0.73 | 3.88 |
| SD | 0.26 | 0.06 | 0.05 | 0.09 | 0.27 | ||
| Range | 0.67 | 0.15 | 0.11 | 0.20 | 0.68 | ||
| Min. | 9.12 | 0.59 | 0.68 | 0.91 | 3.51 | ||
| Max. | 9.80 | 0.73 | 0.79 | 1.11 | 4.19 | ||
| Phymata mystica | |||||||
| males (N=5) | Mean | 8.26 | 0.62 | 0.63 | 1.05 | 0.84 | 3.29 |
| SD | 0.49 | 0.05 | 0.05 | 0.05 | 0.12 | ||
| Range | 1.00 | 0.12 | 0.13 | 0.11 | 0.27 | ||
| Min. | 7.73 | 0.57 | 0.57 | 0.99 | 3.16 | ||
| Max. | 8.72 | 0.69 | 0.70 | 1.10 | 3.43 | ||
| females (N=5) | Mean | 9.19 | 0.62 | 0.67 | 0.88 | 0.68 | 3.57 |
| SD | 0.44 | 0.05 | 0.03 | 0.07 | 0.16 | ||
| Range | 1.05 | 0.12 | 0.07 | 0.15 | 0.34 | ||
| Min. | 8.63 | 0.56 | 0.65 | 0.79 | 3.40 | ||
| Max. | 9.67 | 0.68 | 0.71 | 0.94 | 3.75 | ||
| Phymata obscura | |||||||
| males (N=4) | Mean | 8.35 | 0.57 | 0.63 | 1.05 | 0.87 | 3.23 |
| SD | 0.58 | 0.05 | 0.04 | 0.07 | 0.32 | ||
| Range | 1.31 | 0.11 | 0.10 | 0.18 | 0.76 | ||
| Min. | 7.57 | 0.51 | 0.58 | 0.95 | 2.90 | ||
| Max. | 8.88 | 0.62 | 0.68 | 1.13 | 3.66 | ||
| females (N=4) | Mean | 9.93 | 0.61 | 0.68 | 1.00 | 0.77 | 3.66 |
| SD | 0.33 | 0.04 | 0.04 | 0.08 | 0.35 | ||
| Range | 0.79 | 0.10 | 0.09 | 0.18 | 0.84 | ||
| Min. | 9.52 | 0.56 | 0.65 | 0.89 | 3.31 | ||
| Max. | 10.31 | 0.66 | 0.74 | 1.07 | 4.16 | ||
| Phymata pacifica | |||||||
| males (N=5) | Mean | 6.60 | 0.46 | 0.56 | 0.82 | 0.81 | 2.66 |
| SD | 0.17 | 0.04 | 0.04 | 0.04 | 0.08 | ||
| Range | 0.36 | 0.09 | 0.09 | 0.10 | 0.18 | ||
| Min. | 6.40 | 0.43 | 0.51 | 0.77 | 2.57 | ||
| Max. | 6.76 | 0.52 | 0.60 | 0.87 | 2.75 | ||
| females (N=5) | Mean | 7.85 | 0.52 | 0.63 | 0.70 | 0.61 | 3.17 |
| SD | 0.58 | 0.07 | 0.04 | 0.06 | 0.20 | ||
| Range | 1.59 | 0.18 | 0.08 | 0.13 | 0.44 | ||
| Min. | 7.18 | 0.44 | 0.60 | 0.63 | 2.96 | ||
| Max. | 8.77 | 0.61 | 0.68 | 0.76 | 3.40 | ||
| Phymata paraborica | |||||||
| males (N=8) | Mean | 6.84 | 0.52 | 0.54 | 0.90 | 0.86 | 2.54 |
| SD | 0.18 | 0.03 | 0.03 | 0.05 | 0.09 | ||
| Range | 0.56 | 0.08 | 0.08 | 0.13 | 0.28 | ||
| Min. | 6.59 | 0.48 | 0.50 | 0.84 | 2.38 | ||
| Max. | 7.15 | 0.56 | 0.57 | 0.97 | 2.66 | ||
| females (N=8) | Mean | 7.42 | 0.52 | 0.55 | 0.74 | 0.69 | 2.71 |
| SD | 0.32 | 0.05 | 0.02 | 0.04 | 0.13 | ||
| Range | 1.05 | 0.11 | 0.08 | 0.12 | 0.42 | ||
| Min. | 6.89 | 0.46 | 0.51 | 0.65 | 2.47 | ||
| Max. | 7.94 | 0.56 | 0.59 | 0.78 | 2.88 | ||
| Phymata pennsylvanica | |||||||
| males (N=5) | Mean | 8.19 | 0.56 | 0.59 | 1.32 | 1.15 | 2.92 |
| SD | 0.35 | 0.02 | 0.06 | 0.10 | 0.14 | ||
| Range | 0.94 | 0.05 | 0.16 | 0.25 | 0.34 | ||
| Min. | 7.73 | 0.53 | 0.52 | 1.15 | 2.75 | ||
| Max. | 8.67 | 0.58 | 0.68 | 1.41 | 3.09 | ||
| females (N=5) | Mean | 9.27 | 0.57 | 0.63 | 1.04 | 0.88 | 3.27 |
| SD | 0.22 | 0.05 | 0.09 | 0.04 | 0.13 | ||
| Range | 0.58 | 0.13 | 0.26 | 0.10 | 0.35 | ||
| Min. | 8.97 | 0.51 | 0.50 | 1.01 | 3.16 | ||
| Max. | 9.55 | 0.63 | 0.76 | 1.11 | 3.50 | ||
| Phymata rossi | |||||||
| males (N=3) | Mean | 7.47 | 0.57 | 0.59 | 1.01 | 0.87 | 2.90 |
| SD | 0.77 | 0.07 | 0.04 | 0.03 | 0.29 | ||
| Range | 1.41 | 0.14 | 0.07 | 0.05 | 0.58 | ||
| Min. | 6.59 | 0.52 | 0.57 | 0.98 | 2.61 | ||
| Max. | 8.00 | 0.65 | 0.64 | 1.03 | 3.18 | ||
| females (N=3) | Mean | 8.52 | 0.59 | 0.64 | 0.87 | 0.71 | 3.21 |
| SD | 0.31 | 0.07 | 0.02 | 0.04 | 0.14 | ||
| Range | 0.62 | 0.14 | 0.03 | 0.07 | 0.25 | ||
| Min. | 8.21 | 0.51 | 0.62 | 0.83 | 3.12 | ||
| Max. | 8.82 | 0.65 | 0.66 | 0.90 | 3.38 | ||
| Phymata saileri | |||||||
| males (N=1) | Meas. | 8.01 | 0.58 | 0.56 | 1.04 | 0.91 | 2.86 |
| females (N=1) | Meas. | 8.08 | 0.51 | 0.55 | 0.8 | 0.75 | 3.03 |
| Phymata salicis | |||||||
| males (N=2) | Mean | 7.96 | 0.59 | 0.59 | 0.69 | 0.59 | 2.77 |
| SD | 0.27 | 0.05 | 0.07 | 0.07 | 0.12 | ||
| Range | 0.38 | 0.07 | 0.10 | 0.10 | 0.17 | ||
| Min. | 7.77 | 0.55 | 0.54 | 0.64 | 2.69 | ||
| Max. | 8.15 | 0.63 | 0.64 | 0.74 | 2.85 | ||
| females (N=3) | Mean | 8.34 | 0.54 | 0.59 | 0.59 | 0.52 | 2.84 |
| SD | 0.57 | 0.03 | 0.03 | 0.04 | 0.18 | ||
| Range | 0.99 | 0.06 | 0.06 | 0.07 | 0.33 | ||
| Min. | 8.01 | 0.51 | 0.56 | 0.56 | 2.71 | ||
| Max. | 9.00 | 0.57 | 0.62 | 0.63 | 3.04 | ||
| Phymata stanfordi | |||||||
| males (N=5) | Mean | 7.59 | 0.55 | 0.58 | 0.96 | 0.84 | 2.77 |
| SD | 0.39 | 0.02 | 0.03 | 0.05 | 0.13 | ||
| Range | 1.09 | 0.04 | 0.08 | 0.11 | 0.30 | ||
| Min. | 7.03 | 0.53 | 0.55 | 0.90 | 2.61 | ||
| Max. | 8.13 | 0.57 | 0.63 | 1.01 | 2.92 | ||
| females (N=5) | Mean | 8.40 | 0.62 | 0.65 | 0.81 | 0.65 | 3.07 |
| SD | 0.69 | 0.04 | 0.06 | 0.06 | 0.25 | ||
| Range | 1.60 | 0.09 | 0.16 | 0.11 | 0.64 | ||
| Min. | 7.69 | 0.56 | 0.58 | 0.75 | 2.77 | ||
| Max. | 9.29 | 0.66 | 0.73 | 0.86 | 3.41 | ||
 Figure 1. General morphology of Nearctic erosa group taxa (P. granulosa ♂). Morphological features are abbreviated as follows: aa, anterior angle; af, antennal furrow; apd, anterior pronotal disk; apl, anterior pronotal lobe; bflg, basiflagellomere; cas2–7, connexiva of abdominal segments 2–7; dflg, distiflagellomere; fcx, forecoxa; ffm, forefemur; fp, frontal process; ftb, foretibia; gp, granulation patch; la, lateral angle; lc, longitudinal carina; ln, lateral notch; mspl, mesopleuron; mtpl, metapleuron; op, ocellar process; pa, posterior angle; pd, pedicel; pop, preocellar process; pp, posterior process; ppd, posterior pronotal disk; ppl, posterior pronotal lobe; prpl, propleuron; py, pygophore; sc, scutellum; tb, transverse band of the corium.
Figure 1. General morphology of Nearctic erosa group taxa (P. granulosa ♂). Morphological features are abbreviated as follows: aa, anterior angle; af, antennal furrow; apd, anterior pronotal disk; apl, anterior pronotal lobe; bflg, basiflagellomere; cas2–7, connexiva of abdominal segments 2–7; dflg, distiflagellomere; fcx, forecoxa; ffm, forefemur; fp, frontal process; ftb, foretibia; gp, granulation patch; la, lateral angle; lc, longitudinal carina; ln, lateral notch; mspl, mesopleuron; mtpl, metapleuron; op, ocellar process; pa, posterior angle; pd, pedicel; pop, preocellar process; pp, posterior process; ppd, posterior pronotal disk; ppl, posterior pronotal lobe; prpl, propleuron; py, pygophore; sc, scutellum; tb, transverse band of the corium.
 Figure 2. Pronotal morphology from dorsolateral view (P. fasciata ♂). Morphological features are abbreviated as follows: aa, anterior angle; apd, anterior pronotal disk; apl, anterior pronotal lobe; gp, granulation patch; la, lateral angle; lc, longitudinal carina; ln, lateral notch; pa, posterior angle; pp, posterior process; ppd, posterior pronotal disk; ppl, posterior pronotal lobe; sppl, shoulder of the posterior pronotal lobe; tlc, tubercles of longitudinal carina.
Figure 2. Pronotal morphology from dorsolateral view (P. fasciata ♂). Morphological features are abbreviated as follows: aa, anterior angle; apd, anterior pronotal disk; apl, anterior pronotal lobe; gp, granulation patch; la, lateral angle; lc, longitudinal carina; ln, lateral notch; pa, posterior angle; pp, posterior process; ppd, posterior pronotal disk; ppl, posterior pronotal lobe; sppl, shoulder of the posterior pronotal lobe; tlc, tubercles of longitudinal carina.
![]()
Biological Survey of Canada
Commission biologique du Canada
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}