
The 39 genera of bees found in Canada east of Manitoba are keyed in dichotomous format with all key features illustrated. Habitus photographs of males and females of representatives of all genera are provided. The biology of bees in general, and outlines of the biology of each genus are summarized.
Anthophora furcata (Apidae) Photo by S.A. Marshall.
What are bees?
Bees are one of the most economically important groups of insects as a result of their pollination of agricultural crops (Berenbaum et al., 2006). Although the number of species currently managed for pollination in North America is comparatively small, native wild bees are thought to be responsible for quite a large proportion of the economic benefits attributed to honey bees (Buchmann and Nabhan, 1996). Sometimes, it is not only the number of visits to a flower, but also the diversity of bee species visiting [controlled for total number of insect visits) that determines crop yield (Kremen et al., 2002)]. Wild bees are also crucial for the pollination of most non-crop flowering plants, and thus play an even greater role in most terrestrial ecosystems (Kevan, 2001, 2003; Kevan and Imperatriz-Fonseca, 2002).
Bees are generally considered well known to the general public and thought to be easy to recognize as such. The reality of this is somewhat different, as yellowjackets and hornets (Hymenoptera: Vespidae; Vespinae), hover flies (Diptera: Syrphidae) and other flies are commonly mistaken as being bees. Conversely, many species of bees are superficially more similar to wasps (e.g. Hylaeus Fig 167 and Nomada Figs 176, 177) than are the comparatively hairy and well known honey bees (Fig 144) and bumble bees (Fig 149). The easiest way to know that a specimen at hand is a bee is to locate branched hairs on the body. Most bees are covered in branched hairs, but the comparatively bald groups, such as Hylaeus and Nomada, will require more careful inspection to locate the few and inconspicuous branched hairs. These are generally most easily found on the propodeum or around the pronotal lobe. In Eastern Canada, any insect that is actively collecting pollen onto its hind legs (Figs 149, 18) or the ventral surface of the metasoma is a bee (Fig 172).
Bees are most similar to the apoid wasps (Hymenoptera: Apoidea, families Ampulicidae, Sphecidae and Crabronidae are the ones found in Canada) from which they arose. The branched hair characteristic mentioned above will work to tell these wasps apart from bees in all cases. Other features that differentiate bees from their wasp relatives include the laterally compressed hind basitarsus of bees (which ranges from only weakly compressed, as in Megachile, (Fig 102) to extremely flattened, as in Bombus (Fig 4), and the lack of strongly metallic silvery or golden facial hairs in male bees (many of the related wasps have the lower face covered in strongly reflective hairs).
Canadian Bee Diversity
Published estimates of the number of bee species in Canada range between 520 (Krombein et al., 1979) to over 970 (Danks, 1979). Our current list for Canada has approximately 730 species confirmed in Canada (Sheffield and Packer, unpublished), about 4.0% of the world’s total of approximately 19,000 described species, most of which are found in semi-arid and desertic environments (Michener, 2000). It seems likely that additional surveys and the application of genetic methods to reveal cryptic species (Packer and Taylor, 1997) will increase the number of species in Canada substantially.
Worldwide, bees are divided into nine families (Danforth et al., 2006), three of which (Stenotritidae, Dasypodaidae and Meganomiidae) are not found in Canada (and of these three, only the Dasypodaidae are found anywhere in North America). Of the remaining six families, the Melittidae is rarely found in eastern Canada, while the other five (Megachilidae, Apidae, Andrenidae, Halictidae, and Colletidae) are common throughout Canada except in the far north. The number of bee species in different genera from Canada east of Manitoba is shown in Table 1. Some genera are monotypic, while others contain over 70 species; Andrena and Lasioglossum are the most speciose in our area. The classification adopted here is that of Michener (2000), with the recent changes at the family level suggested by Danforth et al. (2006) and for the tribe Epeolini by Rightmyer (2006). The least stable aspect of the classification adopted is that of the subfamily Panurginae for which the delimitation of genera is under review (J. Ascher, American Museum of Natural History, pers. comm., 2006). In the absence of published phylogenetic analyses, we have opted to follow Michener (2000) for this group, placing bees previously considered to belong to Pseudopanurgus (i.e., Mitchell, 1960) into the genus Protandrena.
Some authors (e.g. Mitchell, 1960; Krombein et al., 1979) prefer a more finely divided classification within the genus Lasioglossum, recognizing the subgenera Lasioglossum, Evylaeus, Sphecodogastra and Dialictus (and other extralimital taxa) at the generic level. Earlier treatments have also treated the Anthophoridae as a separate family (e.g. Krombein et al., 1979; Michener et al., 1994), but it has been shown that the corbiculate bees (honey bees, stingless bees, bumble bees and orchid bees) evolved from within the “Anthophoridae” (Roig-Alsina and Michener, 1993) and so the Apidae as currently understood includes the Anthophoridae of earlier authors.
Bee Biology: Floral Relationships
Bees are digger wasps (Hymenoptera: Apoidea-apoid wasps) that took to a diet of pollen and nectar rather than provisioning animal prey to their offspring. Not surprisingly, bees are known to be almost as old as the flowering plants (Poinar and Danforth, 2006). Most species are mass provisioners, collecting all of the pollen and nectar required for the complete development of an offspring into a single pollen ball before the egg is laid. In such species, there is usually no contact between mother and offspring after oviposition. Bumble bees are the only indigenous bees in Canada in which progressive feeding occurs, the larvae being provided with food by the adults throughout their growth. The only Canadian bee genus to use floral oils as a food source is Macropis which collects oil from Lysimachia (Myrsinaceae).
Some bees are specialists upon one or a few closely related species of flower. Such oligolectic species are generally restricted in their pollen hosts and collect nectar from a wider range of flowers. Examples of oligolectic species include Lasioglossum oenotherae (Stevens), which has scopal hairs adapted for carrying pollen of Oenothera (Onagraceae) [and which can bee seen early in the morning visiting garden varieties such as sundrops (Knerer and MacKay, 1969)], and Melissodes desponsa Smith which collects pollen solely from thistles, Cirsium (Asteraceae). Polylecty, floral host generalization, is more common in the Canadian bee fauna. Individuals of species such as Halictus ligatus Say will collect pollen from more than one family of flowering plant on a single foraging trip.
Cleptoparasitic bees do not collect pollen themselves, but instead lay their eggs on or near the food provisions in the nests of other bees. Cleptoparasitism has arisen independently many times among the bees (Rozen, 2000). The genera which are cleptoparasitic, or include cleptoparasitic species, are shown in Table 1.
Bee Biology: Nests
Most bees nest in the ground, excavating tunnels and constructing brood cells to the side of the tunnel or at the end of a blind burrow. Many ground-nesting bees choose specific soil types for nesting. Sandy soils being chosen by Calliopsis andreniformis Smith and Andrena barbilabris (Kirby) for example, whereas most ground nesting bees are more common in heavier soils. Many species nest in sparsely vegetated ground, Halictus ligatus being one example (Packer and Knerer, 1986), whereas some prefer denser vegetation such that their nests are not so easily detectable. Andrena clarkella (Kirby) is an example of a species that will nest in dense grass. Some bees nest in dense aggregations of up to 300 per square metre, others nest more sparsely.
Another common nesting substrate is in pithy stems, such as those of blackberries and raspberries, Rubus spp. (Rosaceae). Examples of bees that nest in stems include most of the yellow-faced bees Hylaeus, all species of small carpenter bee Ceratina, and many species of the family Megachilidae. Many of the latter also nest in pre-existing cavities in wood, such as holes created by wood-boring beetles, and many of these species accept trap-nests (Krombein, 1967). These bees construct blind, unbranched burrows with the brood cells arranged in a linear series. Abandoned snail shells, or other naturally occurring cavities (even locks) are used by a few species of Osmia (Michener, 2000; O’Toole and Raw, 2004). Some bees, such as Lasioglossum coeruleum (Robertson)(Barrows, 1973), Augochlora pura (Say) (Stockhammer, 1966; Barrows, 1973), Megachile frigida Smith and M. inermis Provancher (Stephen, 1956), excavate burrows in rotting wood, or in harder wood -the large carpenter bee, Xylocopa virginica (L.) being our best example. Bumble bees mostly nest in abandoned rodent burrows, underground or in hollow trees, although some make their nest on the ground (Laverty and Harder, 1988). Honey bees are managed in hives, but feral colonies can be found in hollow trees.
Most bees do not use extraneous materials in constructing their nests. The main exceptions to this are in the family Megachilidae. Species in the genus Megachile line their brood cells with pieces of leaves or petals. The almost perfectly semicircular holes cut in the edges of rose leaves are examples of the work of these bees. Anthidium species line their brood cells with plant hairs, commonly from Stachys (Lamiaceae). Some bees, for example Paranthidium jugatorium (Say), use resin collected from trees or pebbles or mud to line their brood cells and/or to plug nest entrances.
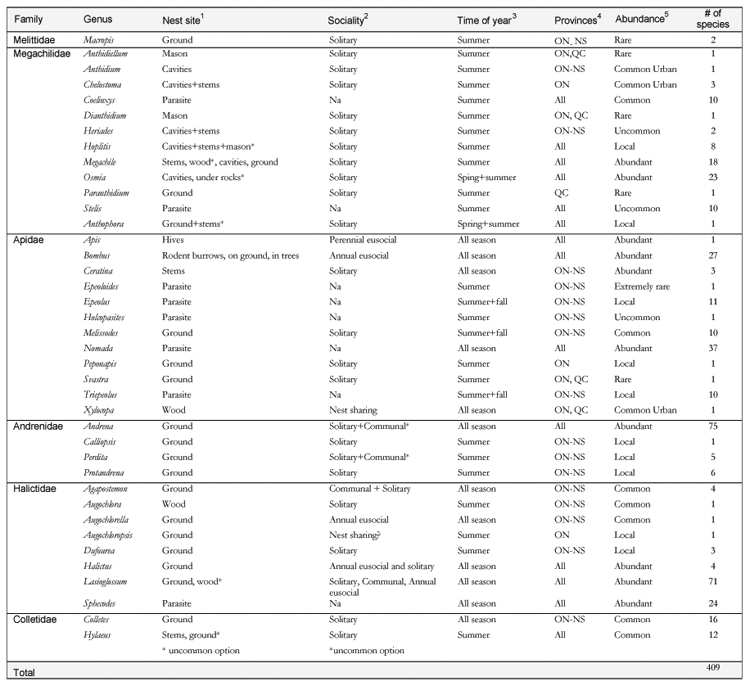
1. Ground: nests made by burrowing into the soil. Mason: nests made on a substrate from resin or mud. Cavities: nests made in naturally occurring cavities such as beetle borings in wood, snail shells, etc. Wood: nests excavated in woody substrates. Stems: nests excavated in pithy stems. Under rocks: (one species) brood cells made under rocks. Hive: the honey bee is the only species that nests in hives, although feral colonies can be found in other hollows and cavities. Rodent burrows: on the ground and in hollow trees; bumble bees nest in these diverse locations. Parasites: make no nests.
2. Solitary: females nest alone. Annual eusocial: overwintered queens start a nest in spring and produce workers before producing males and the next season’s queens. Communal: females share a nest entrance but each constructs her own brood cells and forage for food for their own offspring. Nest sharing: a few females may share a nest, exact details rarely known. Perennial eusocial: the honey bee has colonies that last for many years.
3. Times of year are approximate and will depend upon location. For example, summer bees are unlikely to be found in May or after mid August. All season means that the bees can usually be found from spring to the first frosts.
4. Bees listed as from all provinces can be expected to occur even in northern regions, such as Hudson’s Bay and Labrador. Species listed as ON-NS – or subsets of these provinces, are not generally found far to the north. ON-NS also includes PEI although the bee fauna there has not been well studied yet.
Abundant: easily found almost anywhere. Common: seasonally abundant in most locations. Common Urban: common within urban settings, including gardens. Local: seasonally abundant within specific habitats and locations. Uncommon: seldom encountered but usually widespread. Rare: seldom encountered. Extremely rare: known recently from only a few specimens, possibly extirpated from most of former range.
Most bees are solitary, females constructing a nest and provisioning their brood cells entirely alone. At the opposite extreme are the perennially social colonies of honey bees, comprising one queen and thousands of workers; colonies divide by fission, the old queen leaving in a swarm of approximately half the colony’s workforce to inhabit a new hive (Winston, 1987; O’Toole and Raw, 2004). Honey bees, bumble bees and some sweat bees are eusocial (terminology sensu Michener, 1974): there is a reproductive division of labour between queens that perform all (or at least most) of the reproduction, and their worker daughters. Bumble bees and the eusocial sweat bees have nests started in spring by a single female (some of the sweat bees have nests initiated by several females) which forages to produce the first worker brood. The workers then perform foraging and nest construction activities to produce either more workers (as in bumble bees) or males and next year’s queens (as in the eusocial sweat bees in Canada and late summer workers of bumble bees). Bumble bee colonies can have workforces numbering hundreds of individuals, Canadian sweat bees rarely have more than a dozen workers and some may have an average of less than one worker per nest (Packer, 1990).
Two other forms of social organization are known among Canadian bees: semisociality and communal nesting. Semisocial societies have a reproductive division of labour between females that are of the same generation. The multiple female spring associations in otherwise eusocial sweat bees are examples of this, but some species are known to have at least some colonies that are purely semisocial [e.g. Lasioglossum comagenense (Knerer and Atwood) (Packer et al., 1989)]. Communal bees have no reproductive division of labour, but females share a nest entrance and act as solitary females beneath the ground surface, each constructing and provisioning her own brood cells. The best known example of a communal bee in Canada is Agapostemon virescens (Fabricius), the females of which are readily identified because they are the only eastern Canadian species with the head and mesosoma a brilliant metallic green and the metasoma entirely black with white hair bands. Up to several dozen females may share the same nest in this species (Abrams and Eickwort, 1980).
Bee Conservation
Bees are economically and ecologically important organisms and yet they are more extinction prone than are most other groups of insects (Zayed and Packer, 2005). Oligolectic species may be particularly at risk of extirpation and extinction (Packer et al., 2005; Zayed et al., 2005). There have been dramatic declines in some bumble bee species in recent years, and one species (from the western USA) may have become extinct (Thorp, 2005). One reason why bumble bees are particularly prone to environmental change is that their colonies need large amounts of food over the duration of the entire spring and summer in order to produce males and next year’s queens; solitary bees are not so limited.
Bees need floral and nesting resources. Comparatively few species can use horticultural varieties of flowers for food, whereas many species feed upon plants most people would consider “weeds”. Encouraging bees by planting wildflowers and providing suitable nesting sites would be beneficial. The latter can be done by leaving patches of ground sparsely vegetated and not disposing of dead stems of plants such as raspberry canes. Stem and wood nesting bees can be encouraged by the placement of trap-nests in gardens (Hallett, 2001a, 2001b).
Our key is loosely based upon Michener et al. (1994). We have avoided the use of the most troublesome couplets from that work and have also ensured that the most commonly recognized bee genera (Apis and Bombus) key out early.
We do not present a key to the families of bees for the simple reason that this is based almost entirely on mouthpart characteristics (Michener, 1993) and requires dissections that are not easily performed and are time consuming. Nonetheless, there are some higher level categories that are worth noting.
Bees are divided into short-tongued and long-tongued groups based upon the form of the labial palpi. In short-tongued bees (with few exceptions) the labial palpi are unmodified, whereas in long-tongued bees the basal two segments are considerably elongate, forming a sheath, and the last two segments are very small and at right angles to the basal two. Long-tongued bees evolved from short-tongued ancestors. There are two families of long-tongued bees: the Megachilidae and Apidae (including the Anthophoridae of earlier authors). The remaining families are short-tongued bees.
One short-tongued bee family can be readily identified if the tongue is exposed: the Colletidae are unique in having a glossa that has a concave apex (Fig 79; the few exceptions to this are Australian), so that it is broad at the apex. They use this to brush glandular secretions to make nest cell linings (Torchio et al., 1988). There are only two genera of Colletidae in Canada: Colletes and Hylaeus. Bees with a concave apex to the tongue will either be Colletes (three submarginal cells, lots of pubescence including a scopa on the hind leg in females, face without yellow markings) (Fig 156), or Hylaeus, (two submarginal cells, body almost entirely lacking pubescence, without a scopa, and almost always with yellow/white markings on the face) (Fig 167).
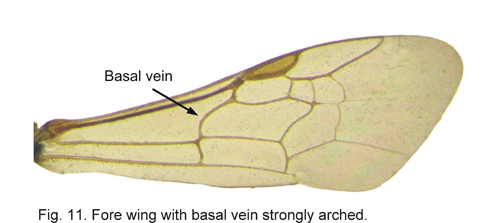
Most Canadian members of the bee family Halictidae can readily be identified by the strongly curved basal vein, most strongly curved towards the base (Fig 11). Lastly, most Canadian non-parasitic members of the bee family Megachilidae can be easily identified by having the scopa entirely on the metasomal sterna of the female (Fig102), other bees with a scopa have it either entirely, or predominantly, on the hind leg (Fig 63).
Species level identification guides, where available, are discussed under each genus. Almost none of these are strictly for the Canadian fauna. We are planning a series of articles that will permit identification of all genera and species of Canadian bees. The Discover Life web site (www.discoverlife.org) has interactive keys to species of most of the genera listed below.
Generic Accounts
In the following we attempt a simple diagnosis for each genus at the beginning of each treatment. We then summarize the known biology of the genus, state its distribution within the area of interest, list available keys and state the number of species in eastern Canada.
1. Agapostemon. (Figs 132, 133). These bees are easily recognized through a combination of two features: bright green head and thorax with a pronounced carina around the posterior surface of the propodeum. The males are also the only bees that are bright green with yellow and black stripes on the metasoma. These are solitary or communal ground-nesting bees that are common in summer in southern regions of our area. Males in particular can often be found flying around Hollyhocks, Alcea spp., and Rose of Sharon, Hibiscus syriaca (both Mavlaceae), flowers in August and September. The four eastern species can be easily separated using Mitchell (1960), though A. radiatus (Say) is now known as A. sericeus (Foster).
2. Andrena. (Figs 134, 135, 136, 137). Andrenidae can be distinguished from other bee families by the paired subantennal sutures. Andrena in our area most commonly have three submarginal cells whereas the other andrenids (Calliopsis, Perdita, Protandrena) have two. Female Andrena can also be readily identified by the facial foveae that are covered in short velvety hairs; all other Canadian bees have hairless facial foveae or, more commonly, no facial foveae at all. These are common solitary ground-nesting bees ranging in size from 5mm to over 15mm, which construct nests in a wide variety of soil types and degrees of vegetation cover. Michener and Rettenmeyer (1956) remains the most detailed study of the nesting biology of any Andrena species. Most species have a black or dark brown integument (a few have red on the metasoma, males of quite a few have pale markings on the lower face). Variable in colour and density of pubescence, most species have dense, mostly brown hairs, while some have bright fuscous pubescence (e.g. A. milwaukeensis Graenicher), others more whitish hairs. Some are comparatively hairless, such as A. integra Smith. They are particularly common visiting willows (Salix, Salicaceae) in spring, but the genus as a whole can be found throughout the spring, summer and autumn. Late summer species are common on goldenrod (Solidago, Asteraceae). These bees can be found throughout eastern Canada, even in the warmer parts of the southern Arctic (Mitchell, 1960). There are approximately 74 species recorded from our area and they can be identified, albeit not easily, using Mitchell (1960), the Discover Life internet site, or using the keys of LaBerge (1967, 1969, 1971, 1973, 1977, 1980, 1986, 1987, 1989), LaBerge and Bouseman (1970), LaBerge and Ribble (1972, 1975), Donovan (1977), Bouseman and LaBerge (1978) and Ribble (1967, 1968).
3. Anthidiellum. (Fig 138). Along with Anthidium, Dianthidium and Paranthidium (discussed below), these bees are easily identified as yellow marked Megachilidae that possess a scopa. Our only species, A. notatum Latreille, which occurs in southern Ontario, can be distinguished from other non-cleptoparasitic Anthidiini by its small size, and the angulate anterior margin of the mesoscutum which overhangs the pronotum. Anthidiellum are summer bees which use resin to construct cells that are attached individually to stems.
4. Anthidium. (Figs 139, 140, 141). These are robust bees with yellow markings on the head and mesosoma, bands of yellow on the metasoma, and no arolia. One indigenous and two introduced Old World species occur in Ontario, one of these, A. manicatum (L.), was also recently reported from Nova Scotia (Hoebeke and Wheeler, 2005). These are the wool carder bees that line their brood cells with hairs from the leaves and stems of plants such as cultivated Lamb’s Ears (Stachys, Lamiaceae). Males are highly territorial and hover around the preferred floral hosts of the females, aggressively pursuing other individuals or other species of bee that visit the plants. Anthidium manicatum is very common in urban gardens in summer where they can be found at mint and other bluish-purple flowers with long corollas, even those kept in flower boxes on balconies of apartment buildings. The species can be distinguished using the keys of Miller et al. (2002) or Romankova (2003a).
5. Anthophora. (Figs 142, 143). These are robust long-tongued bees, with hairless forewing cells and with the outer region of the forewing papillate. Most species nest in the ground, but one, A. terminalis Cresson, excavates nests in pithy stems, and can be found commonly at tomato flowers in urban areas in southern Ontario. These are solitary species. Mitchell (1962) can be used to identify the eastern Canadian species, although A. plumipes (Pallas), a recently introduced European form, is not included in that work and may extend its range into eastern Canada. However, some of the species names in Mitchell (1962) are now considered subspecies or integrades of A. bomboides (Kirby) (see Brooks (1983) for clarification).
6. Apis. (Fig 144). Readily identified by lacking hind tibial spurs, but also by overall appearance, the well known domesticated honey bee, Apis mellifera L., is common throughout the warmer regions of Canada. This introduced species is the only bee in Canada to winter as a colony; all others have a solitary stage in their life history.
7. Augochlora. (Fig 145). These are bright green bees with an acute epistomal lobe. There is only one species of this genus in Canada, A. pura (Say), a bright bluish-green bee that nests solitarily in rotting wood. It can be found through much of the summer and is common in southern Ontario, less so in the Atlantic provinces.
8. Augochlorella. (Figs 146, 147). These are also bright green or bronze bees with a right-angled epistomal lobe, rounded tegula and lacking a carina surrounding the posterior surface of the propodeum. This genus also has only one species in eastern Canada, A. aurata (Smith)(previously known as A. striata (Provancher)). It is a greenish-bronze species, somewhat smaller than A. pura. This eusocial species (Mueller, 1997; Packer, 1990) nests in the ground, forming small turrets at the nest entrance. Augochlorella is common in southern parts of eastern Canada.
9. Augochloropsis. (Fig 148). A third genus of bright green bee with only one species occurs in our area. Augochloropsis metallica (Smith) is a bluish-green species, more robust than either of the preceding species. It can easily be identified by the shape of the tegula. It is a moderately common species in southern Ontario. This species is a ground-nester and it is probable that at least some nests contain multiple females.
10. Bombus. (Fig 149). The common and easily recognized bumble bees are large and furry corbiculate bees with no jugal lobe to the hind wing. All species are eusocial except for the subgenus Psithyrus, which are social parasites that invade host bumble bee nests and replace the queen’s ovipositional activities while relying on her workers to feed their offspring (Fisher, 1987). An additional species not of this subgenus, B. hyperboreus Schönherr, adopts a similar invasion strategy in the far north, but is not a social parasite further south (Richards, 1973). Bumble bees are found throughout our area, and occur even in the arctic where they can forage for 24 hours a day under conditions of continual sunlight. Identification of the 27 eastern species is comparatively easy and mostly based upon details of colouration (Laverty and Harder, 1988).
11. Calliopsis. (Figs 150, 151). The only species in eastern Canada, C. andreniformis Smith, is easily identified by the pattern of white markings on the face of the female and the entirely yellow legs of the male. These bees nest in flat, sandy soil and are locally abundant in suitable areas from southern Ontario to Cape Breton, Nova Scotia, in summer. The biology of this species was discussed in detail by Shinn (1967).
12. Ceratina. (Fig 152). The three species of small carpenter bees found in eastern Canada are all dark metallic blue with ivory markings on the face and/or legs. They nest in dead, pithy stems and, as they overwinter as adults, are among the few Canadian bees that can be collected in midwinter. Although males and females overwinter together, nests that are actively being provisioned house only a single adult female. These bees are active throughout the summer and are found throughout the southern regions of eastern Canada. Mitchell’s (1962) key works well for males, but female identification has recently been improved upon by Rehan et al. (in press), available on the Discover Life website.
13. Chelostoma. (Fig 153). Easily recognized as very narrow and cylindrical Megachilidae, as noted in the key. Three species are known from eastern Canada, two of them introduced from Europe and found in Ontario; the indigenous species, C. philadelphi (Robertson), is known from Ontario and Quebec. One of the introduced species, C. campanularum (Kirby),is a tiny, narrow black bee which commonly visits Campanula (Campanulaceae) flowers in urban areas. These are solitary bees that nest in stems. Buck et al. (2005) provide a key to the species in eastern Canada.
14. Coelioxys. (Figs 154, 155). In our area, these are black bees with bands and/or spots of short dense white hairs. The males have multiple spines at the end of the metasoma, whereas the females’ metasoma ends in a narrow point; these characteristics make this genus easy to recognize. They are found in summer throughout most of eastern Canada and are cleptoparasites that lay eggs in the nests of Megachile species. The ten eastern Canadian species can be identified using Mitchell (1962).
15. Colletes. (Figs156, 157). The S-shaped recurrent vein is diagnostic for this genus, as is the combination of a concave apex of the tongue and three submarginal cells. These bees have a densely pubescent head and mesosoma, usually orange-brown, sometimes dull brown, less commonly blackish or grey. The metasoma is entirely covered in hair in one rare species, C. solidaginis Swenk, but in other species is usually distinctly banded, with the bands at the apex of the terga. These are solitary bees that nest in the ground and line their tubular nests with a cellophane-like secretion that waterproofs the brood cells to prevent both waterlogging from rain and leakage of the rather watery provisions provided for the larvae (Torchio et al., 1988). Different species are active at different times of year such that the genus can be found from early spring (mostly on maple flowers, Acer) to late autumn (mostly on goldenrod, Solidago, Asteraceae). Colletes is common throughout eastern Canada. Romankova (2003b) keyed the Ontario species, but several additional ones have been discovered since, so Mitchell’s (1960) key remains useful for the approximately 17 species.
16. Dianthidium. (Fig 158). One species, D. simile (Cresson), occurs in Ontario.
This is a robust, yellow and black marked bee that can be separated from the other anthidiine genera using the characteristics noted in the key. Dianthidium are mason bees that construct nests of resin and gravel on a surface.
17. Dufourea. (Fig 159). These are small brownish or black bees which have the antennal bases located low on the face, and a scopa which is most strongly developed on the hind tibia. There are three species in southern parts of eastern Canada, each is oligolectic on different plant species. The only moderately common species is D. novaeangliae (Robertson), which forages solely on pickerel weed (Pontederia, Pontederiaceae), an aquatic plant. As such, it is one of the few bees most easily collected while canoeing! The other two species have rarely been seen in our area. These are solitary, ground-nesting bees (Eickwort et al., 1986). Mitchell (1960) identifies the species easily enough.
18. Epeoloides. (Fig 160). This bee is easily identified as a long-tongued cleptoparasite (therefore lacking a scopa) that has no discrete patches or bands of appressed, pale coloured hairs. The one North American species, E. pilosula (Cresson), was thought to be extinct until rediscovered in Nova Scotia (Sheffield et al., 2004). Epeoloides attacks the nests of Macropis and flies in July, but is extremely rare.
19. Epeolus. (Fig 161). These are robust bees with bands and/or spots of white or creamy appressed hairs. The integument is black, sometimes marked with orange or red, especially on the legs. It is only likely to be confused with species of Triepeolus from which it can be separated by the short pseudopygidial area in females and posteriorly convergent margins of the pygidial plate in males. These are cleptoparasitic bees that attack Colletes, and they can often be found flying low over the ground looking for host nests. Mitchell (1962) keyed the 11 species in our area.
20. Halictus. (Fig 162). These are common ground-nesting bees easily identified as halictines by the strongly arched basal vein, strong apical wing venation, and apical hair bands on the metasomal terga. One species (H. confusus Smith) is dull metallic green, the other three are brownish-black. These are socially variable bees. Halictus rubicundus (Christ) is known to be solitary in shorter summer habitats and eusocial in warmer environments (Eickwort et al., 1996) and H. ligatus is one of the best studied social sweat bees (e.g. Richards et al., 1995). Halictus species can be found throughout eastern Canada except in the far north. The four species can be easily identified using Mitchell (1960).
21. Heriades. (Figs 163, 164). These are small, black, coarsely sculptured Megachilidae with a short horizontal portion of the propodeum that is made up of a single row of large pits. They fly in the summer and accept trap-nests. The biology of our most common species, H. carinata Cresson, which occurs as far east as Nova Scotia (Sheffield, 2006), was discussed by Matthews (1965). The three species in our area can be identified with Mitchell (1962).
22. Holcopasites. (Fig 165). The only species in our area, H. calliopsidis (Linsley), is easily identified as a small black and red bee with many spots of white, appressed hairs, giving a checkerboard appearance to the metasoma. It is a cleptoparasite that attacks the nests of Calliopsis andreniformis. It occurs as far east as Nova Scotia (Sheffield, 2006).
23. Hoplitis. (Fig 166). These are small to relatively large, black Megachilidae (a metallic green species occurs in western Canada), similar in appearance to black Osmia, but distinguishable by the elongate parapsidial line on the scutum, and the more elongate metasoma. These summer-flying bees are common throughout our area, some species ranging into the sub-arctic regions (Michener, 2000). Most species can be trap-nested in drilled wood blocks or hollow stems, but a recently recorded species (Buck et al., 2005), H. anthocopoides (Schenck) is a mason which constructs exposed nests out of small pebbles. The eight species in our area (excluding H. anthocopoides) are identifiable using Mitchell (1962).
24. Hylaeus. (Fig 167). Unlike most comparatively glabrous (hairless) species, Hylaeus are not cleptoparasites; rather, they carry pollen and nectar internally and therefore do not possess a scopa. Hylaeus are shiny black bees with white/yellow markings on the face (although females of one species, H. basalis (Smith), are completely black) and with pale banded legs. One species, H. nelumbonis (Robertson), has the basal few metasomal segments red. There are usually a few patches of white hairs on the mesosoma, and on the apicolateral portions of the more basal metasomal terga in some subgenera. Hylaeus are small bees common in the summer, and most nest in stems (and trap-nests) but a few species nest in the ground and, like Colletes, they line their nest with cellophane-like material. The best account of the nesting biology of any Hylaeus species is that of Torchio (1984). Hylaeus in our area can be identified using Mitchell (1960), although H. hyalinatus Smith, an introduced species, was only recently recorded in Ontario (Buck et al., 2005).
25. Lasioglossum. (Figs 168, 169). These are halictines with reduced apical wing venation. The numerous species are classified in four subgenera in eastern Canada: Lasioglossum, Evylaeus, Dialictus and Sphecodogastra. Most nest in the ground, although a few species of the subgenus Dialictus are known to nest in soft wood. This is the most socially diverse genus of bees in the world, with many solitary, a few communal and many eusocial species known. A few species are cleptoparasitic (previously placed in genus or subgenus Paralictus but now considered to be Dialictus – Fig 176). Lasioglossum are very common throughout temperate and boreal Canada, less so in the subarctic, and often make up a large proportion of all specimens collected in surveys (e.g. MacKay and Knerer, 1979; Grixti and Packer, 2006). Larger species (those of the subgenus Lasioglossum) are readily identified using McGinley (1986). The single Eastern Canadian species in the subgenus Sphecodogastra – L. oenotherae (Stevens) is found from Ontario to New Brunswick and is a common early morning visitor to the flowers of evening primroses and sundrops, Oenothera (Onagraceae) (Knerer and MacKay, 1969). The other subgenera, which are typically smaller bees (and many are dull metallic), can be identified, with some difficulty, using Mitchell (1960) although he raised several subgenera to generic rank, incorrectly associated the sexes of several species and made several other mistakes with this group. Ongoing molecular work reveals that there are numerous cryptic species in this difficult group.
26. Macropis. (Figs 170, 171). This is the only genus of Melittidae in Canada. These are uncommon, relatively small, ground-nesting bees that are completely dependent on their floral hosts, Lysimachia (Primulaceae), from which they collect pollen and oils. The oils are used both as a larval food source and as part of the nest cell lining (Cane et al., 1983). Macropis is found in the southern part of our area as far east as Nova Scotia. Mitchell (1960) can be used to separate our two species.
27. Megachile. (Figs 172, 173). The leafcutting bees are readily identified by the combination of metasomal sternal scopa, lack of arolia and lack of yellow integumental markings. All our species use cuttings from leaves or floral petals to line their brood cells which are usually in pre-existing holes in wood (they commonly accept trap-nests), although some species nest in the ground. One species, the alfalfa leafcutter bee, M. rotundata (Fabricius), was accidentally introduced into North America prior to the 1950’s, and is now managed for alfalfa pollination, although it visits many other flowers and has become widely established and is commonly collected. Megachile are common throughout eastern Canada as far north as the sub-arctic zone The 18 species can be identified using Mitchell (1962), although the key uses characteristics of the mandibles, primarily the teeth and cutting edge shape, and identification of specimens with closed mandibles or badly worn teeth, may be difficult.
28. Melissodes. (Figs 174, 175). These are robust, solitary ground-nesting, long-tongued bees. The males have extremely long antennae, in Eastern Canada shared only with Svastra obliqua (Say), which is much less common and rarely collected in our area. Melissodes can be separated by the narrowed anterior portion of the tegula. Mitchell (1962) can be used to separate the ten species in our area; M. rustica (Say) is now known as M. druriella (Kirby).
29. Nomada. (Figs 176, 177). Nomada are wasp-like bees with narrow bodies marked with yellow, orange or red (or combinations of these colours), lacking discrete hair patches and without a strongly curved basal vein. These are mostly cleptoparasites of Andrena and can be easily found flying low above the ground looking for host nests. They are found throughout our area to the arctic circle with different species active at different times of year, although they are most dominant in the early spring. Nomada are often difficult to identify, and numerous species are known from only one sex. Mitchell (1962) can be used to identify the 37 named species in our area and some taxa can be identified using the Discover Life website.
30. Osmia. (Figs 178, 179). These are robust bees, with a dull metallic blue-green integument in most species, a sternal metasomal scopa, and punctiform parapsidal lines. This is the only megachilid genus to overwinter as an adult, and they are thus common in spring and early summer throughout our area, north to the arctic circle. Some individuals in short summer climates skip a year entirely and fly almost two years after their mother laid the egg (Tepedino and Frohlich, 1984). They mostly nest in pre-formed cavities; holes in walls, door locks, and have even been found in stethoscopes in field hospitals! These species commonly accept trap nests. A few species, such as O. inermis (Zetterstedt), build nests under rocks. A variety of extraneous materials are used to line the brood cells, including grass blades, mud, chewed leaves, and gravel. Most of the 23 species can be identified using Mitchell (1962), although additional species have been recorded in our area since that time (Discover Life), and some species are notoriously difficult to identify (Rust, 1974).
31. Paranthidium. (Fig 180). The one uncommon species found in our area, P. jugatorium (Say), can readily be told from all other genera (except Anthidiellum, Anthidium and Dianthidium) by having a sternal scopa, yellow markings on the body, and from these three anthidiine genera by the presence of arolia, a rounded anterior profile to the mesoscutum, and the anterior portion of the mesopleuron which is not separated from the lateral surface by a sharp carina. It nests in sandy soil and lines the brood cells with resin (Evans, 1993).
32. Peponapis. (Fig 181). One species is known in the east, P. pruinosa (Say), the Hoary Squash Bee. This species is most easily identified through its host association: it is oligolectic on curcubits (Cucurbita, Curcubitaceae) and readily found in gardens in southern Ontario. This is a solitary ground-nesting bee found from mid to late summer. Information on this species from Ontario was presented by Kevan et al. (1989).
33. Perdita. (Fig 182). Perdita are small, rather flat bees with body mostly dull metallic and with pale markings on the face and metasoma. These are solitary or communal ground-nesting species and are locally common in southern parts of eastern Canada in the summer and autumn. The nesting biology of the three species found in eastern Canada was reviewed by Michener and Ordway (1963) and Eickwort (1977). The species can be identified using Mitchell (1960).
34. Protandrena. (Figs 183, 184). These are small, black bees with two subantennal sutures and glabrous facial foveae. Species in our area have two submarginal cells. Males have yellow face maculations, including the entire clypeus, though not as extensive as in Calliopsis. These are summer and autumn flying bees, which can often be collected on goldenrod (Solidago) and black-eyed susan (Rudbeckia, Asteraceae). Mitchell (1960) can be used to identify the six species in our area (as the genus Pseudopanurgus).
35. Sphecodes. (Fig 185). This is our only genus of cleptoparasitic halictines (strongly arched basal vein), which are also shiny, usually coarsely sculptured and with the metasoma largely red (but entirely black in males of a few species). These bees mostly attack the nests of other halictines. Common throughout our area south of the arctic circle. There are 24 species recorded from our area, and they can be identified with difficulty using Mitchell (1960). One reason for their taxonomic complexity is that species attacking a single eusocial host with strong size dimorphism between queens and workers, or a species attacking multiple hosts of different sizes, will exhibit marked size variation according to which caste or species of host the parasite female laid eggs on.
36. Stelis. (Figs 186, 187).Identifiable as cleptoparasitic Megachilidae with yellow markings on the metasoma. These bees attack Megachilidae, primarily Heriades and Hoplitis, but possibly Osmia and Chelostoma as well. The ten species can be identified using Mitchell (1962).
37. Svastra. (Figs 188, 189). One species is found in our area: S. obliqua (Say). It can be separated from other bees, except Melissodes, by the enormously long antennae in the male, and from Melissodes (and all other bees) by the spatulate hairs on some parts of the body (see the key for details). It is rare, known in eastern Canada only from southern Ontario and Quebec.
38. Triepeolus. (Fig 190). Readily identified from all other genera except Epeolus as a long-tongued cleptoparasitic bee with appressed patches/bands of white or yellowish hairs, giving a strongly patterned appearance. It is separated from Epeolus by the longer psuedopygidial area in females and sinuate lateral margin to the pygidial plate in males. These are cleptoparasites, mostly of Melissodes. Ten species are found in southern parts of eastern Canada. They can be identified using Mitchell (1962) or Rightmyer (2006).
39. Xylocopa. (Fig 191). One species is found in eastern Canada: Xylocopa virginica (L.). These large carpenter bees nest in wood and can be pests of outdoor wooden structures. Nest sharing by females is known and they are comparatively long-lived as adults. These bees are becoming increasingly common in southern Ontario and seem to be spreading north.
![]()
Biological Survey of Canada
Commission biologique du Canada
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}