
Cantharidae, also known as soldier beetles, is a group of very common beetles commonly found as adults on foliage and in flowers, where they feed on small insects, nectar and pollens. Some species are involved in the natural control of aphid populations that infest plants, shrubs and trees. This synopsis of the Cantharidae of Eastern Canada and Northeastern United States covers 114 species among the following genera: Atalantycha Kazantsev (3 species), Cantharis Linnaeus (8), Pacificanthia Kazantsev (2), Rhagonycha Eschscholtz (31), Rhaxonycha Motschulsky (2), Dichelotarsus Motschulsky (17), Podabrus Dejean (22), Ditemnus LeConte (2), Polemius LeConte (4), Silis Charpentier (3), Malthinus Latreille (1), Malthodes Kiesenwetter (14), Trypherus LeConte (2) and Chauliognathus Hentz (2). All species are described morphologically along with data on distribution, seasonal occurrence, habitats (when known), number of specimens examined and, for most species, proportion of females and males. Some species not recorded in Canada yet but living in bordering States in similar habitats are also included.
Several taxonomic changes are suggested in this publication. Cantharis brevicollis (LeC.) is removed from synonymy with Cantharis tuberculata (LeC.). However, Cantharis impressa (LeC.), listed as a distinct species in McNamara (1991), is now a synonym of Cantharis tuberculata (LeC.). Cantharis curtisi (Kby.)is transferred to Pacificanthia. Dichelotarsus has been removed from the synonymy of Podabrus. The following species are transferred from Podabrus to Dichelotarsus: cinctipennis (LeC.), deceptus (Brown), extremus (LeC.), fumiganus (Green), heteronychus (Fall), laevicollis (Kby.), limbellus (LeC.), pattoni (LeC.), perplexus (Brown), piniphilus (Eschs.), probus (Fall), puberulus (LeC.), punctatus (LeC.), puncticollis (Kby.), secretus (Brown), and simplex (Couper). One Holarctic species, Dichelotarsus flavimanus, is recorded for the first time in North America. Podabrus moestus Fall, described from California, is now a synonym of D. flavimanus Motsch, which is an Holarctic species.
The following 24 species are recorded for the first time in Canada east of the Rocky Mountains: Cantharis alticola (LeC.)(MB, SK, AB), C. brevicollis (LeC.)(QC, ON), C. livida (L.) (QC), C. transmarine scopa (LeC.)(SK, AB), Rhagonycha angulata (Say) (ON), R. greeni (Fall) (ON), R. hirticula (Green) (ON), R. lineola (F.) (ON), R. nigrohumeralis (Green) (ON), R. parvicollis (Green) (NS, QC, ON), R. proxima (Green)(NB, QC, ON), R. tenuis (Green) (NB, QC, ON), R. umbrina (Green)(NB, QC, ON), Dichelotarsus cinctipennis (LeC.) (ON, QC), D. flavimanus Motsch. (NB, QC, ON, MB, AB, BC, NU, NT, YK), Podabrus basillaris (Say) (QC, ON), P. dreisbachi Green (ON), P. longicornis Fall (NB, QC, MB), Silis spathulata LeC. (QC, ON), Polemius limbatus LeC. (ON), Polemius repandus LeC. (NB, QC), Malthodes concavus LeC. (QC, ON), Malthodes werneri Fend. (QC) and Trypherus latipennis (Germ.) (ON).
Excluding the species mentioned previously, there is at least one new provincial record for each of the following 42 species: Atalantycha bilineata (Say) (NB), Cantharis tuberculata (LeC.) (NS, NB, ON, MB), Rhagonycha imbecillis (LeC.) (NS, NB, QC), R. luteicollis (Germ.) (NB, MB), R. mandibularis (Kby.) (NB, ON), R. mollis mollis (Fall) (QC), R. nanula (LeC.) (QC, ON), R. oriflava (LeC.) (QC, SK), R. recta (Melsh.) (NB, MB, SK), R. scitula (Say) (ON), R. septentrionis (Green) (NB, QC, SK, AB), R. sylvatica (Green) (QC), R. tantilla (Say) (QC, ON, MB, AB), R. walshi (LeC.) (QC, ON, SK), Rhaxonycha carolina (F.) (NB), Dichelotarsus deceptus (Brown) (QC), D. extremus (LeC.) (QC), D. limbellus (LeC.) (NS, ON), D. pattoni (LeC.) (NS, ON), D. perplexus (Brown) (QC, ON, AB, BC, YK), D. piniphilus (Eschs.) (NS, NB, ON, SK), D. probus (Fall) (NB), D. puberulus (LeC.) (SK, AB), D. punctatus (LeC.) (ON), D. puncticollis (Kby.) (ON, SK, BC), D. secretus (Brown) (NL), Podabrus brevicollis Fall (QC), P. flavicollis LeC. (QC), P. frosti Green(NS, QC), P. modestus (Say) (NB), P. nothoides LeC. (NB), P. planulus Green (NB), P. punctulatus LeC. (MB), P. pygmaeus Green (ON), Ditemnus latilobus (Blatchl.) (QC, ON), Malthodes fragilis (LeC.) (YK), M. fuliginosus LeC. (ON), M. medioccidens Fend. (QC), M. niger (LeC.) (NS, ON), M. parvulus (LeC.) (MB, SK, AB, BC), M. similis Fend. (QC) and Chauliognathus pensylvanicus (DeGeer) (MB).
To summarize, we mention one new record for Newfoundland and Labrador, eight new records for Nova Scotia, 19 for New Brunswick, 31 for Quebec, 34 for Ontario, ten for Manitoba, ten for Saskatchewan, eight for Alberta, four for British Columbia, one for the Northwest Territories, one for Nunavut and three for Yukon.
Résumé
Les Cantharidés, connus sous le nom de Téléphores en Europe (et de Cantharides au Canada), constituent un groupe de Coléoptères très communs vivant sur le feuillage et les fleurs au stade adulte et se nourrissant d’insectes variés, de nectar et de pollens. Certaines espèces sont impliquées dans le contrôle naturel des populations de pucerons qui infestent les plantes, les arbustes et les arbres. Ce synopsis des Cantharidés de l’Est du Canada et du Nord-Est des États-Unis couvre 114 espèces appartenant aux genres suivants : Atalantycha Kazantsev (3 species), Cantharis Linnaeus (8), Pacificanthia Kazantsev (2), Rhagonycha Eschscholtz (31), Rhaxonycha Motschulsky (2), Dichelotarsus Motschulsky (17), Podabrus Dejean (22), Ditemnus LeConte (2), Polemius LeConte (4), Silis Charpentier (3), Malthinus Latreille (1), Malthodes Kiesenwetter (14), Trypherus LeConte (2) et Chauliognathus Hentz (2). Toutes les espèces sont décrites morphologiquement avec des données de répartition géographique, d’occurrence saisonnière, d’habitats (lorsque connus), du nombre de spécimens examinés et, pour la majorité des espèces, de la proportion des femelles et des mâles. Quelques espèces, non encore découvertes au Canada, mais vivant dans des États voisins et dans des habitats semblables, sont aussi décrites dans cette publication.
Parmi les changements taxonomiques présentés dans cette publication, mentionnons que Cantharis brevicollis (LeC.) a été exclu de la synonymie de Cantharis tuberculata (LeC.). Cependant, Cantharis impressa (LeC.), mentionnée comme une espèce distincte dans McNamara (1991), est maintenant un synonyme de Cantharis tuberculata (LeC.). Cantharis curtisi appartient maintenant au genre Pacificanthia. Dichelotarsus est exclu de la synonymie de Podabrus. Les espèces suivantes ont été transférées de Podabrus à Dichelotarsus : D. cinctipennis (LeC.), D. deceptus (Brown), D. extremus (LeC.), D. fumiganus (Green), D. heteronychus (Fall), D. laevicollis (Kby.), D. limbellus (LeC.), D. pattoni (LeC.), D. perplexus (Brown), D. piniphilus (Eschs.), D. probus (Fall), D. puberulus (LeC.), D. punctatus (LeC.), D. puncticollis (Kby.), D. secretus (Brown) et D. simplex (Couper). Une espèce holarctique, Dichelotarsus flavimanus Motschulsky, est mentionnée pour la première fois en Amérique du Nord. Podabrus moestus Fall, décrite à partir de spécimens provenant de la Californie, est maintenant un synonyme de D. flavimanus.
Les 24 espèces suivantes sont mentionnées pour la première fois au Canada à l’est des Rocheuses : Cantharis alticola (LeC.)(MB, SK, AB), C. brevicollis (LeC.)(QC, ON), C. livida (L.) (QC), C. transmarine scopa (LeC.)(SK, AB), Rhagonycha angulata (Say) (ON), R. greeni (Fall) (ON), R. hirticula (Green) (ON), R. lineola (F.) (ON), R. nigrohumeralis (Green) (ON), R. parvicollis (Green) (NS, QC, ON), R. proxima (Green)(NB, QC, ON), R. tenuis (Green) (NB, QC, ON), R. umbrina (Green)(NB, QC, ON), Dichelotarsus cinctipennis (LeC.) (ON, QC), D. flavimanus Motsch. (NB, QC, ON, MB, AB, BC, NU, NT, YK), Podabrus basillaris (Say) (QC, ON), P. dreisbachi Green (ON), P. longicornis Fall (NB, QC, MB), Silis spathulata LeC. (QC, ON), Polemius limbatus LeC. (ON), Polemius repandus LeC. (NB, QC), Malthodes concavus LeC. (QC, ON), Malthodes werneri Fend. (QC) et Trypherus latipennis (ON).
Excluant les espèces citées précédemment, il y a plus de 42 espèces nouvellement mentionnées dans au moins une province : Atalantycha bilineata (NB), Cantharis tuberculata (LeC.) (NS, NB, ON, MB), Rhagonycha imbecillis (LeC.) (NS, NB, QC), R. luteicollis (Germ.) (NB, MB), R. mandibularis (Kby.) (NB, ON), R. mollis mollis (Fall) (QC), R. nanula (LeC.) (QC, ON), R. oriflava (LeC.) (QC, SK), R. recta (Melsh.) (NB, MB, SK), R. scitula (Say) (ON), R. septentrionis (Green) (NB, QC, SK, AB), R. sylvatica (Green) (QC), R. tantilla (Say) (QC, ON, MB, AB), R. walshi (LeC.) (QC, ON, SK), Rhaxonycha carolina (NB), Dichelotarsus deceptus (Brown) (QC), D. extremus (LeC.) (QC), D. limbellus (LeC.) (NS, ON), D. pattoni (LeC.) (NS, ON), D. perplexus (Brown) (QC, ON, AB, BC, YK), D. piniphilus (Eschs.) (NS, NB, ON, SK), D. probus (Fall) (NB), D. puberulus (LeC.) (SK, AB), D. punctatus (LeC.) (ON), D. puncticollis (Kby.) (ON, SK, BC), D. secretus (Brown) (NL), Podabrus brevicollis Fall (QC), P. flavicollis LeC. (QC), P. frosti Green(NS, QC), P. modestus (Say) (NB), P. nothoides LeC. (NB), P. planulus Green (NB), P. punctulatus LeC. (MB), P. pygmaeus Green (ON), Ditemnus latilobus (Blatchl.) (QC, ON), Malthodes fragilis (LeC.) (YK), M. fuliginosus LeC. (ON), M. medioccidens Fend. (QC), M. niger (LeC.) (NS, ON), M. parvulus (LeC.) (MB, SK, AB, BC), M. similis Fend. (QC) et Chauliognathus pensylvanicus (DeGeer) (MB).
Pour conclure, nous rapportons une nouvelle mention pour Terre-Neuve et Labrador, huit pour la Nouvelle-Écosse, 19 pour le Nouveau-Brunswick, 31 pour le Québec, 34 pour l’Ontario, dix pour le Manitoba, dix pour la Saskatchewan, huit pour l’Alberta, quatre pour la Colombie-Britannique, une pour les Territoires du Nord-Ouest, une pour le Nunavut et trois pour le Yukon.

Chauliognathus pensylvanicus - Photo by Peter Cristofono
Cantharidae, also known as soldier beetles or cantharids, are common beetles often found as adults on foliage and flowers, where they feed on insects, nectar and pollen. Several species have been reported to prey on aphid populations infesting plants, shrubs and trees (Way and Banks 1968; Berthiaume et al., 2001; Day et al. 2006), but they can also attack other groups of insects. Indeed, Chauliognathus pulchellus (Macleay) feed on eggs of a chrysomelid beetle, Chrysophtharta bimaculata (Olivier), in Tasmanian forests (Mensah and Madden 1994). Nevertheless, the life history and ecology of most species of Cantharidae are poorly known. For instance, it is known that larvae are mostly carnivorous in the soil (Fender 1973), but their specific prey remain unknown. Using an electrophoretic approach, Traugott (2003) showed that larvae of three Cantharis species fed on earthworms and on dipterous and lepidopterous larvae but much remains to be done to highlight the ecological role of cantharids in terrestrial ecosystems.
Most species emerge in spring or summer and adults are short lived. They are among the most active flying beetles (Ramsdale 2002). They are predominantly active during the day but they may stop activity if temperature becomes too hot (Ramsdale 2002). Many species are also active at night as they were collected in much higher numbers in pitlight traps than in passive pitfall traps (Hébert et al., 2000). Being more exposed to predators on the surface of vegetation, they have developed an effective system of chemical defense (Dettner, 1987). Adults and larvae possess paired tergal glands that secrete repulsive compounds that serve to reduce their palatability to predators (Ramsdale, 2002).
This review was prepared because cantharids were frequently captured in our research projects aimed to determine the impact of forestry practices on beetle diversity in Canadian forests. We hope that the publication of a modern tool for identifying cantharid species will enhance interest on their study and thus increase our knowledge on their life history and ecology. This is strongly needed to help interpreting results in biodiversity projects and to improve our understanding of ecosystem functioning.
Cantharids are easily distinguished from other beetles by their soft elytra and their head which is not completely concealed from above. A total of 473 species belonging to 25 genera have been described so far in North America north of Mexico. McNamara (1991) listed 126 species in Canada and suggested that probably 25 undescribed or unrecorded species remained to be added to our fauna.
The taxonomy of Cantharidae is relatively well known in Canada and the United States due to the extensive work of Kenneth M. Fender, Dorothy McKey-Fender and John W. Green. Cantharis,the nominate genus, was described by Linnaeus in 1758 with C. fusca as the type species. Cantharis rufa and C. livida (both introduced in North America) were among the first species of the family to be named by Linnaeus. Thomas Say described 12 cantharids under the name Cantharis between 1823 and 1835 (Say 1823, 1825, 1835). During much of the 19th century, the name «Cantharidae» was used for beetles now included in the family Meloidae. The first nearctic revision of the family was done by LeConte in 1851 who also described many species of Telephorus, which were then assigned to the subfamily Telephorinae of the larger family Lampyridae, but which are now included in Cantharis and related genera. LeConte described many species of Podabrus between 1850 and 1881 (LeConte 1850, 1866a, 1866b, 1881), as well as some species of Malthinus and Malthodes (sometimes as Malthinus). Fall (1928) revised Podabrus and described many new species. All the primary types of LeConte and Fall are at the Museum of Comparative Zoology, were photographed and are available at http://insects.oeb.harvard.edu/mcz/findrecords.php.
A major revision of Rhagonycha, as a subgenus of Cantharis, was done by Green (1940). Brown (1940) described three new species of arctic Podabrus (now belonging to Dichelotarsus) and Green (1947, 1948) added further contributions concerning that genus. McKey-Fender (1950) partly revised Cantharis (now belonging to Atalantycha and Rhaxonycha). However, since those publications, the former genus Cantharis has been divided into five different genera: Atalantycha (Kazantsev 2005), Cantharis, Pacificanthia (Kazantsev 2001), Rhagonycha (Fender 1971) and Rhaxonycha (Ramsdale 2002). More recently, Dichelotarsus, described by Motschulsky in 1860, and long considered as a subgenus of Podabrus in Europe, was restored by Kasantsev (1992). Fender (1951) produced a major contribution concerning Malthodes that included all eastern North American species. Trypherus, then newly discovered in Canada,was revised by Fender (1960). Finally, Green (1966) revised Silis (including the species now in Ditemnus), described many new species, but none in our area. Downie and Arnett (1996) provided keys and brief descriptions of northeastern North American species.
Cantharids are widely distributed in North America north of Mexico. Atalantycha, which contains three species, is found only in eastern North America. Rhagonycha, Podabrus, Dichelotarsus and Cantharis are widely distributed, with the former two genera being more diversified in the east and Dichelotarsus more highly diversified west of the Rocky Mountains. Silis and Ditemnus are more diversified in southwestern North America. The large genus Malthodes is mostly diversified in western and southern North America, with relatively few species in the northeast. Members of Chauliognathus are mainly found in the south with only two species in Canada.
Cantharidae is found in a wide variety of habitats. Over the last 20 years, we sampled hundreds of forest stands throughout the province of Quebec. Based on these studies and on specimen labels from various collections, we can categorize Podabrus and Dichelotarsus as general forest dwellers. Atalantycha and Silis were mostly found in hardwood forests and Pacificanthia in conifer forests. Rhagonycha is more diversified in hardwood and pine forests, marshes and shrubby areas. Cantharis and Chauliognathus are common in grasslands and forb fields. Malthodes is common in mixed and conifer forests in eastern North America.
Cantharid seasonality can also be overviewed on the basis of specimen labels. Atalantycha can be found early in spring from April to May in Canada. Most cantharids (Podabrus, Dichelotarsus, Rhagonycha, Malthodes) are very active in June, with populations decreasing in July. However, Silis, Ditemnus and Polemius are predominant in July and Chauliognathus pensylvanicus is mostly seen in August and September.
This publication covers 114 described species that are found in eastern Canada and northeastern United States, as defined by Downie and Arnett (1995). The region covered includes Newfoundland west to Ontario, south to New Jersey, Pennsylvania, Ohio, Indiana, Illinois and Wisconsin. All known species found in Manitoba and Minnesota are also included as they bordered Ontario.
All the types of LeConte and Fall were verified by the senior author. However, four species described by Miskimen (1956) from Ohio, three belonging to Rhagonycha and one to Dichelotarsus,have not been verified with the types and have not been incorporated into the key. Two of these species were described from a single specimen and in one case from the female only. Kazantsev’s (2004) list of Cantharidae of the former USSR was checked in order to verify if some of our Arctic species could be synonymised with some Palearctic species. Four species were suspected as potential synonyms and specimens of two Palearctic species were borrowed from European museums to compare with our Nearctic species (the two other species could not be found).
This publication is available in PDF format as well as an interactive identification key online. This digital key will lead to images that can be selected in order to progress to the final diagnosis of the species. For each species, a brief description is provided as well as images of the habitus and genitalia (for Malthini and few other groups) and often field photographs. Species variations, comparison with similar species, habitat, ecological data, seasonality, and distribution maps are also given. Photographs of habitus or morphological structures were taken with a Nikon Digit-like Camera DMX 1200F through a Nikon SMZ 1500 stereoscopic microscope. All photos were processed through a Proscan II system by Prior, which combines all images taken at different layers into one focused image. Each body part (head, thorax, elytra, antennae, legs) of each species was photographed individually and then joined together using Adobe Photoshop software.
Though species are well defined taxonomically, existing keys for many genera can be used only for identifying males. Some characters used in the previously published keys were complex and confusing, making identification of many species difficult. As an example, Green (1940) only used claws of the first pair of legs of males in Rhagonycha (as Cantharis) to separate many species because of the great variability of this character. However, for many species, according to our data, males usually represent about 20% of the adult population and not more than 1% in Rhagonycha fraxini. In such cases, the available identification keys are almost useless for separating most specimens. For that reason, we decided to use the claws of the metathoracic legs of both sexes, which are similar and less variable but, combined with other characters, like the front margin of the clypeus, elytral pilosity and pronotum shape, help to separate all species in both sexes. We hope that this publication will help students, amateurs, technicians and entomologists to easily identify most species, including female specimens.
Most cantharid specimens in the Canadian National Collection, in Ottawa, were examined. Many specimens from Quebec came from the biodiversity project led by Christian Hébert at the Laurentian Forestry Centre. Steve Marshall from the University of Guelph provided many specimens and some ecological data from Ontario. Reggie Webster also provided specimens from New Brunswick to complete the picture. Claude Chantal and Michel Racine gave a substantial number of specimens from Quebec for identification or confirmation.
Live pictures were used with permission of authors from the Bug Guide site (http://bugguide.net/node/view/118/bgpage). Steve Marshall also provided a substantial amount of images for this publication.
Both sexes are described together and not separately as in Green (1940). When there is sexual dimorphism, females are described first because they are usually more numerous than males in the majority of species. Length is measured from the anterior margin of the clypeus to the apex of the elytra. When measuring, you need to consider that the apex of the elytra is often folded downward and this folded area must be extrapolated in the calculation. All characters necessary to identify the species (diagnosis) are bolded in the text.
Each part of the beetle is described in this sequence: head (including eyes, clypeus, mandibles, palpi, antennae), thorax (pronotum, sternum, legs) and abdomen (elytra, ventrites), describing coloration first, then morphometry and other structural characters. Eyes/head width ratio is calculated by dividing the distance between eyes (frons width) at the level of maximum head width by the head width (from dorsal view) and subtracting the result from 1: 1-(FW/HW) where FW=frons width and HW=head width. This ratio of eyes/head width is calculated for both females and males because males have larger eyes than females. The clypeus can be notched, biarcuate, sinuate or rectilinear. Antennae/body length ratio is calculated for many genera. The third and second antennomeres length ratio (A3/A2), both for females and males, is also recorded.
Pronotum color, width/length ratio, margin and tubercle shapes, surface porosity, punctures and pubescence are noted. Width/length ratios of pronotum go from subquadrate (1:1) to 2X as wide as long. Lateral margins can be arcuate, sinuate, rectilinear, subparallel, incised or irregularly modified (in males of Silis and Ditemnus). Anterior angles can be arcuate or angulate. Many species have two distinct tubercles near basal margin, often prominent and with distinct characteristic shape (Podabrus, Dichelotarsus). Pronotum can be glossy, subglossy or opaque (with distinct microsculpture) with strong, moderate or sparse punctures. Pubescence is usually moderately long and sparse but sometimes very short and dense. Coloration of prosternum, meso-, metaventrite and scutellum are mentioned. Leg coloration is described followed by the character state of the tarsal claws of both females and males which can be simple, cleft or toothed at base.
Elytra coloration, length/width ratio and the elytra/pronotum length ratio are given. In many cases, the elytra are folded on the sides (costal margins) and at the apex, which make morphometric measurement difficult and modify its shape. As an example, Rhagonycha fraxini has elytra with their lateral margins divergent, but because the elytra are folded on the sides, the lateral margins will look subparallel. Some other species have the elytra separately arcuate at the apices but because they are folded apically, they appear truncate. The elytra need to be examined laterally and posteriorly to verify if the foldings give a biased appearance to the shape. The elytra can have the lateral margins subparallel, divergent, or rarely convergent. The surface can be scabrous or rugose with the pubescence dense, moderate or sparse, erect or/and prostrate. In some cases, mostly in Malthini, median lobe of aedeagus and subgenital plate of the male are described. In Malthini, males have the aedeagus exerted in dried and mounted specimens. In Cantharini and Podabrini, median lobe of aedeagus of most species are illustrated but not described.
Finally, colour variations are noted in many species, which can easily lead us to confuse several species. Variations in body shape are essentially due to specimen preparation artifacts because of their soft body. All similar species related to the one described are mentioned with characters that can help to separate them, starting with the most similar species. These characters are usually mentioned in the key.
Distributions are given from eastern Canada and northeastern USA to the northwest down to the southeast and then to the southwest, including the range outside the geographic scope of this work. Distribution maps are given for each species and are overlaid on Canadian ecozones and US ecoregion maps, as defined below. However, for simplication, some ecozones/ecoregions were grouped. All names come from ArcGIS map software from ESRI. In Canada, the ecological framework comes from Agriculture and Agri-Food Canada and Environment Canada (http://sis.agr.gc.ca/cansis/publications/manuals/1996/cad-map.jpg). In the United-States, it is a modification of Bailey’s ecoregions from USDA Forest Service (http://www.fs.fed.us/rm/ecoregions/products/map-ecoregions-united-states/#). Legend for ecozones/ecoregions outside the scope of this work is not described below.
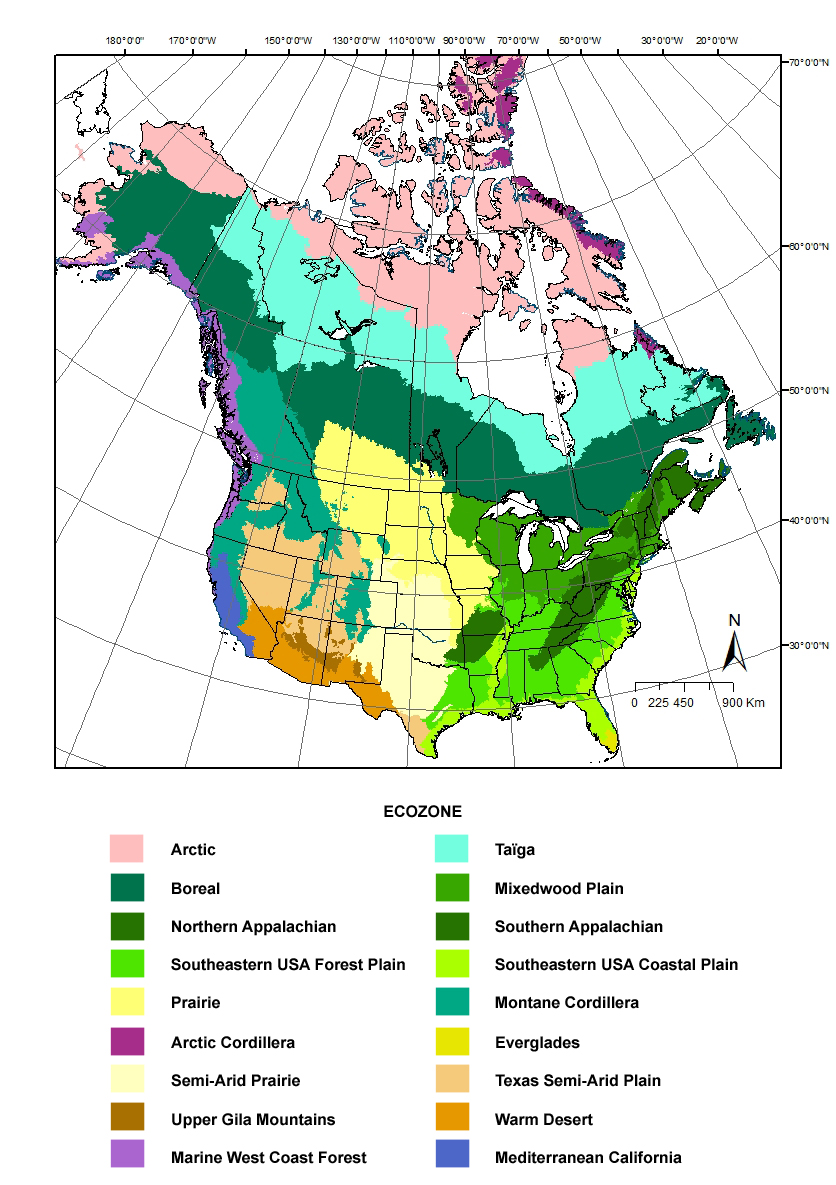
Arctic zone, in northern Canada (including most Arctic Ocean islands) to Alaska, from coast to coast, covered mostly by toundra.
Taïga zone, also called Subarctic and sometime Hudsonian zone, bordering south of the Arctic zone, from coast to coast, covered mostly by open Black Spruce forest growing sparsely on lichen.
Boreal zone, also called Canadian zone, mostly covered by dense Black Spruce forest, more dominated by Balsam Fir in the south, extending nearly coast to coast.
Mixedwood Plain zone, mostly covered by deciduous forests predominating by maple-beech, but also mixed forests with White Pine and Red Oak in wet areas, along the St-Lawrence River and the Maritime provinces and states. This area is mostly covered by agricultural lands.
Northern Appalachian zone, including Atlantic Maritime zone of Canada, mountain chain extending from eastern Quebec, New Brunswick south to western Massachusett, Connecticut and Adirondack Mountains of New York, a southern extension of the Boreal zone, dominated by spruce-fir and maple-beech forests.
Southern Appalachian zone, (also called Alleghanian zone), mountain chains extending from Pennsylvania to north Georgia and Alabama, including Ozark mountain of Missouri and Arkansas, southern extension of the northern Appalachian and the Mixedwood zones, covered by maple-beech forests but with higher proportion of pine and oak-hickory forests. Spruce-fir forests can be found in highest mountains.
Southeastern USA Forest Plain zone (also called Carolinian zone), extending south of the Mixedwood and the Southern Appalachian zones, from New Jersey to north Florida west to Illinois, Arkansas and Texas, predominating by pine and oak-hickory forests.
Southeastern USA Coastal Plain zone, (including the Mississipi Alluvial and Southeastern USA Coastal Plain, the Texas-Louisiana Coastal Plain, bordering the Atlantic and the Gulf of Mexico seashore south of the Southeastern Forest Plain zone, from New Jersey to Florida west to the Mississipi valley and Texas, predominating by vast pine forests and evergreen hardwood forests predominated by oak.
Prairie zone, excluding South Central Semi-Arid Prairie, west of the Boreal, Mixedwood Plain and Southeastern USA Forest Plain zones, from southern Manitoba to Alberta south to Missouri, Kansas, Nebraska, and Wyoming, predominated by grasslands and meadows.
Montane Cordillera, including the cordillera of western USA, from British Columbia and western Alberta south to Colorado, New Mexico, Utah, Idaho and California, predominated by ponderosa and lodgepole pines, very diversified area containing many endemic species, but also extending range of few eastern species.
Habitats, when known, are reported for each species. However, as ecological data were not recorded systematically by collectors in the past, habitats remain unknown for about 20% of the species in the area covered. However, after the 1992 Earth Summit in Rio de Janeiro, Brazil, a structured biodiversity research program was launched at Laurentian Forestry Centre of the Canadian Forest Service and habitats of many species, mostly in eastern Canada, began to be better known. Our program allowed increasing knowledge for species living in spruce, fir and maple forests. Moreover, specimen label data were useful for improving knowledge of other species living in grasslands, open shrubby fields and orchards which are relatively well known. However, though some species have been recorded in these habitats, oak and pine forests still need to be more intensively investigated, as well as freshwater and coastal marshes and bogs.
Seasonality is based on data from labeled specimens collected mostly in eastern Canada. For species with high number of specimens, the information is much more detailed, including specification about peak of activity. Finally, the number of specimens examined is given to indicate the relative abundance of the species in collections as well as the proportion of females and males to indicate the relative scarcity of males in some species. Females usually outnumber males in collections.
| Family CANTHARIDAE Imhoff, 1856 (Soldier beetles) |
|||||||||||
| Subfamily CANTHARINAE Inhoff, 1856 | |||||||||||
| Tribe Cantharini Inhoff, 1856 | |||||||||||
| Genus ATALANTYCHA Kazantsev, 2005 Key to species: Kazantsev (2005) |
|||||||||||
| A. bilineata (Say, 1823) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| signaticollis (Dejean, 1833) | |||||||||||
| marginalis (Gemminger and Harold, 1869) | |||||||||||
| A. dentigera (LeConte, 1851) | – | – | – | – | – | – | ON | QC | – | – | – |
| A. neglecta (Fall, 1919) | – | – | – | – | – | – | ON | QC | – | – | – |
| Genus CANTHARIS Linnaeus, 1758 | |||||||||||
| C. alticola (LeConte, 1881) | – | – | BC | AB | SK | MB | – | – | – | – | – |
| C. aneba McKey–Fender, 1951 | – | – | – | AB | SK | MB | ON | – | – | – | – |
| C. brevicollis (LeConte, 1866) | – | – | – | – | – | – | ON | QC | – | – | – |
| tuberculata (LeConte, 1851) in part | |||||||||||
| C. lecontei Fall, 1936 | – | – | – | – | – | – | ON | – | – | – | – |
| collaris (LeConte, 1852) | |||||||||||
| torquata (Gemminger, 1870) | |||||||||||
| C. livida Linnaeus, 1758† | – | – | – | – | – | – | ON | QC | NB | – | – |
| C. rufa Linnaeus, 1758† | – | – | – | – | – | – | ON | QC | NB | NS | NF |
| C. transmarina scopa (LeConte, 1866) | – | – | BC | AB | SK | – | – | – | – | – | – |
| C. tuberculata (LeConte, 1851) | – | – | – | – | – | MB | ON | QC | NB | NS | – |
| impressa (LeConte, 1851) | |||||||||||
| armiger (Couper, 1865) | |||||||||||
| Genus PACIFICANTHIA Kazantsev, 2002 Key to species: Kazantsev (2002) |
|||||||||||
| P. curtisi (Kirby, 1837) | YT | NT | BC | AB | SK | MB | ON | QC | NB | – | NF |
| samouellii (Kirby, 1837) | |||||||||||
| P. rotundicollis (Say, 1825) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| Genus RHAGONYCHA Eschscholtz, 1830 Key to Nearctic species: Green (1940) |
|||||||||||
| R. angulata (Say, 1823) | – | – | – | – | – | – | ON | – | – | – | – |
| R. cartwrighti (Green, 1940) | – | – | – | – | – | – | – | – | – | – | – |
| R. dichroa (LeConte, 1851) | – | – | – | – | – | – | ON | – | – | – | – |
| flavipes (LeConte, 1851) | |||||||||||
| gilvipes (Gemminger, 1870) | |||||||||||
| R. excavata (LeConte, 1881) | – | – | BC | – | – | MB | ON | QC | – | NS | – |
| R. fraxini (Say, 1823) | – | – | BC | AB | – | MB | ON | QC | NB | NS | NF |
| ater (Kirby, 1837) | |||||||||||
| binodula Mannerheim, 1846 | |||||||||||
| nigritus (LeConte, 1850) | |||||||||||
| R. fulva (Scopoli, 1763)† | – | – | BC | – | – | – | ON | QC | NB | NS | – |
| R. greeni (Fall, 1936) | – | – | – | – | – | – | ON | – | NB | – | – |
| R. heterodoxa (Green, 1940) | – | – | – | – | – | – | – | – | – | – | – |
| R. hirticula (Green, 1940) | – | – | – | – | – | – | ON | QC | – | – | – |
| R. imbecillis (LeConte, 1851) | – | – | – | – | – | MB | ON | QC | NB | – | – |
| albolineatus (Blatchley, 1817) | |||||||||||
| R. lineola (Fabricius, 1792) | – | – | – | – | – | – | ON | – | – | – | – |
| parallela (Say, 1825) | |||||||||||
| sayi (LeConte, 1851) | |||||||||||
| R. longula (LeConte, 1852) | – | – | – | – | – | – | ON | – | – | – | – |
| R. luteicollis (Germar, 1824) | – | – | – | – | – | MB | ON | – | NB | – | – |
| cinctellus (LeConte, 1851) | |||||||||||
| R. mandibularis (Kirby, 1837) | YT | NT | BC | AB | SK | MB | ON | QC | NB | – | NF |
| nigritulus (LeConte, 1881) | |||||||||||
| R. mollis mollis (Fall, 1936) | – | – | – | – | SK | – | ON | QC | NB | NS | NF |
| nigriceps (LeConte, 1850) | |||||||||||
| pusillus (LeConte, 1881) | |||||||||||
| mimus (Fall, 1936) | |||||||||||
| R. nanula (LeConte, 1881) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| R. nigrohumeralis (Green, 1940) | – | – | – | – | – | – | ON | – | – | – | – |
| R. oriflava (LeConte, 1874) | – | – | – | – | SK | – | ON | QC | – | – | – |
| R. parvicollis (Green, 1940) | – | – | – | – | – | – | ON | QC | – | NS | – |
| R. proxima (Green, 1940) | – | – | – | – | – | – | ON | QC | NB | – | – |
| R. recta (Melsheimer, 1846) | – | – | BC | AB | SK | MB | ON | QC | NB | NS | NF |
| rufipes (Say, 1823) | |||||||||||
| pusillus (LeConte, 1851) | |||||||||||
| R. scitula (Say, 1825) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| R. septentrionis (Green, 1941) | – | – | – | AB | SK | MB | ON | QC | NB | – | – |
| R. sylvatica (Green, 1941) | – | – | – | – | – | – | ON | QC | – | – | – |
| R. tantilla (LeConte, 1881) | – | – | – | AB | SK | MB | ON | QC | – | – | – |
| pusio (LeConte, 1881) | |||||||||||
| R. tenuis (Green, 1940) | – | – | – | – | – | – | ON | QC | NB | – | – |
| R. triangulifera (Green, 1940) | – | – | – | – | – | – | – | – | – | – | – |
| R. umbrina (Green, 1940) | – | – | – | – | – | – | ON | QC | NB | – | – |
| R. vestigialis (Green, 1940) | – | – | – | – | – | – | – | – | – | – | – |
| R. vilis (LeConte, 1851) | – | – | – | – | – | – | ON | QC | NB | – | – |
| R. walshi (LeConte, 1881) | – | – | – | – | SK | MB | ON | QC | – | – | – |
| Genus RHAXONYCHA Motschulsky, 1860 Key to Nearctic species: McKey-Fender (1950) |
|||||||||||
| R. bilobata (McKey-Fender, 1941) | – | – | – | – | – | MB | ON | QC | NB | – | – |
| R. carolina (Fabricius, 1801) | – | – | – | – | – | – | ON | QC | NB | – | – |
| jactatus (Say, 1825) | |||||||||||
| Tribe Podabrini Gistel, 1856 | |||||||||||
| Genus DICHELOTARSUS Motschulsky, 1833 | |||||||||||
| D. cinctipennis (LeConte, 1866) | – | – | – | – | – | – | ON | QC | NB | – | NF |
| D. deceptus (W.J. Brown, 1940) | YT | NT | BC | – | SK | MB | – | QC | – | – | – |
| D. extremus (LeConte, 1881) | YT | NT | BC | AB | – | MB | ON | QC | – | – | NF |
| D. flavimanus Motschulsky, 1860 | YT | NT | BC | AB | – | MB | ON | QC | NB | – | – |
| impressa (LeConte, 1851) | |||||||||||
| moestus (Fall, 1928) | |||||||||||
| heteronychus (not Fall, 1928) in part | |||||||||||
| D. fumiganus (Green, 1948) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| D. heteronychus (Fall, 1927) | YT | NT | BC | AB | SK | MB | ON | QC | – | – | – |
| D. laevicollis (Kirby, 1837) | YT | NT | BC | AB | SK | MB | ON | QC | NB | – | NF |
| D. limbellus (LeConte, 1881) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| D. pattoni LeConte, (1866) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| D. perplexus (W.J. Brown, 1940) | YT | NT | BC | AB | – | MB | ON | QC | – | – | – |
| D. piniphilus (Eschscholtz, 1830) | YT | NT | BC | AB | SK | MB | ON | QC | NB | NS | NF |
| D. probus (Fall, 1927) | – | – | – | – | – | – | – | QC | NB | – | – |
| D. puberulus (LeConte, 1850) | – | – | – | AB | SK | MB | ON | QC | NB | NS | NF |
| D. punctatus (LeConte, 1850) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| D. puncticollis (Kirby, 1837) | – | – | BC | AB | SK | MB | ON | QC | NB | – | – |
| marginellus (LeConte, 1850) | |||||||||||
| D. secretus (W.J. Brown, 1940) | – | – | – | AB | – | – | – | QC | – | – | NF |
| D. simplex (Couper, 1865) | – | – | – | – | – | MB | ON | QC | NB | NS | – |
| Genus PODABRUS Dejean, 1833 | |||||||||||
| P. appendiculatus Fall, 1927 | – | – | – | – | – | – | ON | QC | – | – | – |
| P. basillaris (Say, 1823) | – | – | – | – | – | – | ON | QC | – | – | – |
| pennsylvanicus (Melsheimer, 1851) | |||||||||||
| discoidea LeConte, 1851 | |||||||||||
| P. brevicollis Fall, 1927 | – | – | – | – | – | – | ON | QC | – | – | – |
| P. brimleyi Green, 1947 | – | – | – | – | – | – | – | – | – | – | – |
| P. brunnicollis (Fabricius, 1801) | – | – | – | – | – | – | – | – | – | – | – |
| puncticollis LeConte, 1851 | |||||||||||
| poricollis LeConte, 1851 | |||||||||||
| P. diadema (Fabricius, 1798) | – | – | – | AB | – | – | ON | QC | NB | NS | – |
| P. dreisbachi Green, 1947 | – | – | – | – | – | – | ON | – | – | – | – |
| P. fayi LeConte, 1866 | – | – | – | – | – | – | ON | – | – | – | – |
| P. flavicollis LeConte, 1851 | – | – | – | – | – | – | ON | QC | – | – | – |
| P. frater LeConte, 1851 | – | – | – | – | – | – | ON | – | – | – | – |
| quadricollis (Motschulsky, 1859) | |||||||||||
| P. frosti Fender, 1946 | – | – | – | – | – | – | ON | QC | NB | NS | – |
| P. intrusus Green, 1947 | – | – | – | – | – | – | ON | QC | NB | NS | NF |
| P. longicornis Fall, 1928 | – | – | – | – | – | MB | – | QC | NB | – | – |
| P. modestus (Say, 1823) | – | – | – | – | – | – | ON | QC | NB | NS | NF |
| P. nothoides LeConte, 1881 | – | – | – | – | – | MB | ON | QC | NB | – | – |
| P. planulus Green, 1947 | – | – | – | – | – | MB | ON | QC | NB | NS | – |
| P. protensus LeConte, 1866 | – | – | – | – | – | – | ON | – | – | – | – |
| P. punctulatus LeConte, 1859 | – | – | – | – | – | MB | ON | QC | – | – | – |
| P. pygmaeus Green, 1948 | – | – | – | – | – | – | ON | QC | – | – | – |
| P. rugosulus LeConte, 1850 | – | – | – | AB | – | MB | ON | QC | NB | NS | – |
| parvicollis (Motschulsky, 1859) | |||||||||||
| P. tomentosus (Say, 1825) | – | – | – | – | – | – | ON | – | – | – | – |
| rufiolus (Melsheimer, 1846) | |||||||||||
| cinereipennis Motschulsky, 1859 | |||||||||||
| rubricollis Gemminger and Harold, 1869 | |||||||||||
| P. tricostatus (Say, 1835) | – | – | – | – | – | – | ON | QC | – | – | – |
| bennetti (Kirby, 1837) | |||||||||||
| atripes (Motschulsky, 1859) | |||||||||||
| Subfamily SILINAE Mulsant, 1862 | |||||||||||
| Tribe Silini Mulsant, 1862 | |||||||||||
| Genus DITEMNUS LeConte, 1861 | |||||||||||
| D. bidentatus (Say, 1825) | – | – | – | – | – | – | ON | QC | – | NS | – |
| lepida (Dejean, 1837) | |||||||||||
| D. latilobus Blatchley, 1910 | – | – | – | AB | – | MB | ON | QC | – | – | – |
| Genus POLEMIUS LeConte, 1851 | |||||||||||
| P. canadensis W.J. Brown, 1940 | – | – | – | – | – | – | ON | QC | NB | NS | – |
| P. laticornis (Say, 1825) | – | – | – | – | – | – | ON | QC | NB | – | – |
| invalida (Say, 1835) | |||||||||||
| dubius (Melsheimer, 1846) | |||||||||||
| incisus LeConte, 1851 | |||||||||||
| undulatus (LeConte, 1851) | |||||||||||
| P. limbatus LeConte, 1851 | – | – | – | – | – | – | ON | – | – | – | – |
| P. repandus LeConte, 1881 | – | – | – | – | – | – | – | QC | NB | – | – |
| Genus SILIS Charpentier, 1825 Key to Nearctic species: Green (1966) |
|||||||||||
| S. difficilis difficilis LeConte, 1850 | YT | NT | BC | AB | SK | MB | – | QC | – | – | – |
| carbo Van Dyke, 1918 | |||||||||||
| occidens Green, 1966 | |||||||||||
| S. percomis (Say, 1835) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| longicornis LeConte, 1850 | |||||||||||
| curtus (LeConte, 1850) | |||||||||||
| S. spathulata LeConte, 1881 | – | – | – | – | – | – | ON | QC | – | – | – |
| Subfamily MALTHININAE Kiesenwetter, 1852 | |||||||||||
| Tribe Malthinini Kiesenwetter, 1852 | |||||||||||
| Genus MALTHINUS Latreille, 1806 | |||||||||||
| M. occipitalis LeConte, 1851 | – | – | – | – | – | – | ON | QC | – | – | – |
| atripennis LeConte, 1881 | |||||||||||
| woodruffi Wittmer, 1980 | |||||||||||
| Tribe Malthodini Böving & Craighead, 1930 | |||||||||||
| Genus MALTHODES Kiesenwetter, 1852 | |||||||||||
| M. arcifer LeConte, 1881 | – | – | – | – | – | – | – | – | – | – | – |
| M. captiosus LeConte, 1881 | – | – | – | – | – | – | – | – | – | – | – |
| hirsutotibialis Fender, 1845 | |||||||||||
| M. concavus (LeConte, 1851) | – | – | – | – | – | – | ON | QC | – | – | – |
| M. curvatus LeConte, 1881 | – | – | – | – | – | – | – | – | – | – | – |
| M. fragilis (LeConte, 1851) | YK | – | – | – | – | – | ON | QC | NB | NS | – |
| transversus (LeConte, 1851) | |||||||||||
| M. fuliginosus fuliginosus LeConte, 1866 | – | – | – | – | – | – | ON | QC | NB | NS | – |
| M. medioccidens Fender, 1951 | – | – | – | – | – | – | ON | QC | – | – | – |
| M. megapygus Fender, 1951 | – | – | – | – | – | – | – | – | – | – | – |
| M. niger (LeConte, 1851) | – | – | – | – | – | – | ON | QC | NB | NS | – |
| M. parvulus (LeConte, 1851) | – | – | BC | AB | SK | MB | ON | QC | NB | NS | – |
| M. rectus LeConte, 1881 | – | – | – | – | – | – | – | – | – | – | – |
| M. similis Fender, 1951 | – | – | – | – | – | MB | ON | QC | NB | NS | – |
| M. spado LeConte, 1866 | – | – | – | – | – | – | – | – | – | – | – |
| M. werneri Fender, 1951 | – | – | – | – | – | – | – | QC | – | – | – |
| Tribe Ichthyurini Champion, 1915 | |||||||||||
| Genus TRYPHERUS LeConte, 1851 | |||||||||||
| T. frisoni Fender, 1960 | – | – | – | – | – | – | ON | – | – | – | – |
| T. latipennis (Germar, 1824) | – | – | – | – | – | – | ON | – | – | – | – |
| marginalis (Say, 1824) | |||||||||||
| nigriceps (Dejean, 1837) | |||||||||||
| Subfamily CHAULIOGNATHINAE LeConte, 1861 | |||||||||||
| Tribe Chauliognathini LeConte, 1861 | |||||||||||
| Genus CHAULIOGNATHUS Hentz, 1830 | |||||||||||
| C. marginatus (Fabricius, 1775) | – | – | – | – | – | – | ON | – | – | – | – |
| americanus (Forster, 1771) | |||||||||||
| ligatus (Say, 1825) | |||||||||||
| philadelphicus (Dejean, 1833) | |||||||||||
| marginipennis (Laporte, 1840) | |||||||||||
| hentzii LeConte, 1851 | |||||||||||
| C. pensylvanicus (DeGeer, 1774) | – | – | – | – | – | MB | ON | QC | NB | – | – |
| bimaculatus (Fabricius, 1781) | |||||||||||
(Based on Ramsdale (2002))
Description: Length 1.0 to 15 mm. Body elongate, often parallel-sided, mostly flattened, soft, with lightly sclerotized cuticle, especially on elytra and abdomen. Body usually with moderately dense, short, depressed pubescence. Head completely or partially visible from above. Eyes usually entire, moderately to strongly protruding; ocelli absent. Antennae elongate, pubescent, with eleven antennomeres; antennal insertions exposed from above, moderately to widely separate in most species. Labrum free, membranous, often concealed beneath clypeus. Mandibles elongate, curved with acute apex. Maxillae with densely setose galea and lacinia in most species; maxillary palpi 4 segmented; labial palpi 3-segmented.
Pronotum with lateral margins usually entire; base as wide as or narrower than base of elytra. Scutellum well developed and visible. Prothoracic coxae conical, prominent, projecting below prosternum, contiguous or nearly so; prothoracic coxal cavity absent; prothoracic trochanter exposed. Mesothoracic coxae contiguous or nearly so; cavities open laterally. Metaventrite with lateral margins sinuate; metathoracic coxae flattened, contiguous or narrowly separated. Legs elongated and slender in most species with paired tibial spurs (except in Chauliognathini). Tarsal formula 5-5-5; fourth tarsomere expanded and ventrally bilobed.
Elytra soft, more or less parallel-sided or divergent, loosely fitted to pterothorax and sternites. Metathoracic wings usually well developed.
Abdomen with 7 (females) or 8 (males) freely moveable sternites with complete sutures; tergites 1-8 with paired lateral glandular pores. Male genitalia of trilobed type; tegmen composed of the phallobase (basal piece) and lateral lobes (parameres). Median lobe membranous in most species, with eversible internal sac, highly variable in structure and often ornamented with asperities. Additional sclerotized genitalic processes arising from interior of tegmen. Female genitalia with coxites and paraprocts well developed in most species.
![]()
Biological Survey of Canada
Commission biologique du Canada
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}